OS MARSUPIAIS SÃO PLACENTÁRIOS MODIFICADOS
 |
| HOMOLOGIES - palatines and pterygoids |
Os marsupiais "são placentários modificados" e isso muda muita coisa. Muda o olhar da genética, mudam as referencias, pois os grupos externos agora são os Xenarthra e os Pholidota. Muita coisa será compreendida sob esse novo olhar.
Estou aqui tentando estabelecer homologias entre "répteis" e mamíferos. Tentando compreender a evolução dos ossos palatinos e pterigoides. Tanto o contato entre esses 2 ossos, como também a "proximidade" entre o pterigoide e o quadrado. Isso nos permitirá compreender a evolução do palato e sua relação com o aparato auditivo.
Podemos observar em Varanus uma separação entre os palatinos e pterigoides. Como cada lado do crânios se desenvolve separadamente no embrião e depois se juntam, quando ocorre PEDOMORFOSE podemos ter resultantes desse tipo (um basicrânio aberto)
A fenestração observada no palato de Didelphis (opossum), que é comum nos Didelphidae, também pode ser vista como uma interrupção no processo de desenvolvimento embrionário (uma pedomorfose).
Os pterigoides que nos répteis estão muito próximos do quadrado, nos marsupiais estão afastados da região auditiva. Mas em Tachyglossus (equidna) e nos pangolins estão em contato com o aparato auditivo. Lembrando que o osso quadrado nos mamíferos sofreu uma redução e deu origem a bigorna (incus), um dos ossículos do ouvido médio.
PALATAL EVOLUTION IN MAMMALS

I am focusing on this board in the palatal region (yellow), in the pterygoids (orange) and in the auditory region (blue and green). The skulls were calibrated by the anterior zygomatic arch, as shown by the markings. The only exception is Ornithorhynchus which was calibrated by length with only Tachyglossus.
1) The comparisons showed a gradation in the placement of the palatine. Located well back in the Monotremata (as happens in some reptiles), to a central location with respect to the length of the skull. The arrangement of palatal shown in Tachyglossus is quite different from all others resembles a reptilian conformation, especially its relationship with the pterygoids which are also obviously reptilian.
2) The pterygoids in Ornithorhynchus are also large and reptilian but apparently in mammalian course. These bones are present in Xenarthra and more reduced derivatives in Metatheria.
3) The region of ear ossicles (auditory) varies from a disposition facing the ventral side, and without a tympanic bulla in the Monotremata, or tympanic ring formed by the merger of ectotympanic + endotympanic as in Xenarthra (Pretheria) and that is probably the same one found in the Pholidota and Tubulidentata, to a bulla almost completely closed by advancing alisfenóide and facing the side, as in the case of Metatheria.
The analysis of these three regions points to a similar trend to those already suggested by other previous analysis I posted. Reaffirms the basal position of Monotremata, in accordance with the traditional and current view of the group. In Didelphis (Metatheria) and Orycteropus (Eutheria), the posterior region of the palate shows close phylogenetic relationships among these groups, putting marsupials within the taxon Placentalia. These relationships have already been mentioned by me earlier, with respect to the ear ossicles. This has agrupament like sisters groups Pholidota first, and then Pretheria more external (as seen in the figure above)
Sinodelphys is an extinct mammal from the Early Cretaceous. To date, it is the oldest metatherian fossil known, estimated to be 125 million years old. It was discovered and described in 2003 in rocks of the Yixian Formation in Liaoning Province, China, by a team of scientists including Zhe-Xi Luo and John Wible. Wikipedia
The skeleton, undoubtedly represents a plesiomorphic form of marsupial, very similar in some respects to Caenolestes, in particular the tarsal elements. We know nothing about their internal anatomy, if they had a placenta, similar to the Eutheria, or if they made nests for their newborns, as Ornithorhynchus, or Xenarthra and Pangolins. We know that Sinodelphys was already differentiated from other Theria and in line with current marsupials. We know that Juramaia is considered a Eutheria and lived in the middle Jurassic (163 million to 174 million years ago), is strong evidence that placentals, with 8 post-canine teeth as Cabassous lived 40 million years before Sinodelphys. Given the strong evidence presented by me in the table above, and other fantastic posts that I made on FLICKR, everything indicates that marsupials are modified placentals .
Apparently this group of mammals dispersed to the southern continents, which at that time were in a colder region than that inhabited by Sinodelphys.
† Glossopteris is a fossil plant of the Permian, which has been used as a reference to characterize the union of the southern continents (Gondwana), became extinct in the late Permian. Its distribution was apparently circum-polar, which indicates that in the Jurassic, South America probably was in a temperate zone of the southern hemisphere, and throughout South America had a colder climate than today.
I want to propose here, that both Tachyglossus, as current marsupials developed a strategy to keep her cubs away from the ground next to the body as do the penguins, which hatch their eggs on the feet, close to the body. In this case Sinodelphys despite being in line with the current marsupials, was still a nest-maker, and that changes in reproduction occurred only in the southern continents, where the temperature was a problem, apart from scarcity of food, which also favored animals could move with their young.
The ordering, and therefore the polarization of these characters presented above is based on evolving evidence. This order is justified by synapomorphies. Although not from organized matrices and not being result of computer analysis, the requirements for a good phylogenetic proposal are fulfilled perfectly.
Marsupial Evolution and Biogeography
I am presenting here an update of the idea that was being propagated on the evolution and biogeography of Marsupials. We'll have to forget forever the ideas concerning the existence of a Antarctica bridge, that served as passage to the dispersion of Marsupials to Australia.
Who is watching my discoveries and details about the evolution of the Earth, you know that the Antarctic Peninsula did not belong to the continent as always thought. This peninsula stood in the middle of South America and Africa, was probably not in direct contact with Antarctica.
Also as we have seen, the Andes had training also different from what you thought. I divided this story of the genesis of the Andes in three steps. Two of them can even be simultaneous, but undoubtedly the central region is the newest. This central region was formed in the Cretaceous with the opening of the South Pacific. The relationship of the Andes with the emergence and evolution of Marsupials became much more plausible and coherent. The oldest fossils found so far, show us an Asiatic origin for the Marsupials. The discovery of Sinodelphys szalayi Luo et al., 2003.
- LUO, Z.-X.; JI, Q.; WIBLE, J. R.; YUAN, C.-X. (2003). An Early Cretaceous Tribosphenic Mammal and Metatherian Evolution. Science 302: 1934-1939.) was the first step toward change to a new insight into the origin and dispersal of Marsupials.
- LUO, Z.-X.; JI, Q.; WIBLE, J. R.; YUAN, C.-X. (2003). An Early Cretaceous Tribosphenic Mammal and Metatherian Evolution. Science 302: 1934-1939.) was the first step toward change to a new insight into the origin and dispersal of Marsupials.
My studies, about continental drift, show that Asia was in direct contact with the North America and South America before the opening of the Pacific Ocean. The oldest fossils of marsupials are from the Lower Cretaceous. At this time the North Pacific was already open, but parts of Asia were still in direct contact with Australia and probably had intermittent contact during the Cretaceous. Australia in turn was in contact with South America during the entire Cretaceous.
The biogeographical evolution of non-asian Marsupials appear to have begun in the northern Andes, where we find the Caenolestidae, the most conservative Marsupials of the planet (representing by Caenolestes fuliginosus, the most conservative species). As I have shown, in the plates that I published on this album of the FLICKR, these marsupials still retain a plesiomorphic heel structure very similar to that of Sinodelphys, much more conservative than any other living or fossil marsupial outside Asia.
The Australian and South America Marsupials (except Caenolestidae) share a derived type of heel, with a more triangular Astragalus which can be seen in this figure and compared to the Caenolestidae. Among Caenolestidae known, "Caenolestes fuliginosus" can be considered the most conservative of them all. Their geographical distribution is in the northern Andes, roughly between the Bay of Guayaquil and the far north the Cordillera. Exactly the ancient region of the Andes. Also another interesting distribution is to Dromiciops gliroides exactly in the region by me named Andes II. Dromiciops as we have seen is the sister group of Didelphimorphia retaining some plesiomorphic characteristics already modified in others, such as a non-stagged 3 lower incisor,(see i3 stagged in the figure, one important synapomorphy of Didelphimorphia).
See "Hershkovitz, P. 1992a. Ankle bones: The Chilean opossum Dromiciops gliroides Thomas, and marsupial phylogeny. Bonner Zool. Beiträge 43 (2): 181-213".
See "Hershkovitz, P. 1992a. Ankle bones: The Chilean opossum Dromiciops gliroides Thomas, and marsupial phylogeny. Bonner Zool. Beiträge 43 (2): 181-213".
Finally the Didelphimorphia, representing the South American Marsupials (except Caenolestidae and Dromiciops) plus Australians, originated in this new formation, I called Andes III, during the Cretaceous. The Marsupials fossils from North America and Europe, considering its features regarding heel, probably originated from the northern populations of South America because they do not share the synapomorphies cited for Didelphimorphia. Hershkovitz however pointed a stagged incisor 3 in some fossils of the northern hemisphere, which means that at least part of that marsupials colonized North America were more closely related to Didelphimorfia. Hershkovitz also included the Caenolestidae in Didelphimorphia however the great development of i2 in these Marsupials makes that conclusion seem dubious.
In the picture "Caenolestes fuliginosus" (the most conservative species) above left; "Marmosa murina" right; "Eudromicia caudata" lower left, the astragalus is from another species "Distoechurus pennatus" and also down below "Dromiciops gliroides".
 |
| Na foto Caenolestes fuliginosus (a espécie mais conservadora do grupo) acima à esquerda; Marmosa murina à direita; Eudromicia caudata em baixo à esquerda, o astrágalo é de outra espécie Distoechurus pennatus e também em baixo a seguir Dromiciops gliroides. |
Estou apresentando aqui uma atualização da ideia que vinha sendo propagada sobre a evolução e a biogeografia dos Marsupiais. Teremos que esquecer para sempre as ideias referentes à existência de uma ponte Antártica que funcionou como passagem para a dispersão dos Marsupiais até a Austrália.
Quem está acompanhando minhas descobertas e detalhes sobre a evolução do Planeta Terra, sabe que a Península Antártica não pertencia ao continente como sempre se pensou. Essa península se destacou do meio da América do Sul e da África, provavelmente nem estava em contato direto com a Antártida.
Além disso como vimos, os Andes tiveram uma formação também diferente daquela que se pensava. Dividi essa história da gênese dos Andes em três etapas. Duas delas podem até ser simultâneas, mas sem dúvida a região central é bem mais nova. Essa região central se formou no Cretáceo com a abertura do Pacífico sul. A relação dos Andes com o aparecimento e a evolução dos Marsupiais se tornou muito mais plausível e coerente. Os fósseis mais antigos até agora encontrados nos mostram uma origem asiática para os Marsupiais. A descoberta de Sinodelphys szalayi Luo et al., 2003
- LUO, Z.-X.; JI, Q.; WIBLE, J. R.; YUAN, C.-X. (2003). An Early Cretaceous Tribosphenic Mammal and Metatherian Evolution. Science 302: 1934-1939.) foi o primeiro passo em direção a mudança para uma nova visão sobre a origem e dispersão dos Marsupiais.
- LUO, Z.-X.; JI, Q.; WIBLE, J. R.; YUAN, C.-X. (2003). An Early Cretaceous Tribosphenic Mammal and Metatherian Evolution. Science 302: 1934-1939.) foi o primeiro passo em direção a mudança para uma nova visão sobre a origem e dispersão dos Marsupiais.
Meus estudos sobre Deriva Continental mostram que a Ásia estava em contato direto coma América da Norte e a América do Sul antes da abertura do Oceano Pacífico. Os fósseis mais antigos de marsupiais são do Cretáceo Inferior. Nessa época o Pacífico norte já estava aberto, mas certas regiões da Ásia ainda se encontravam em contato direto com a Austrália e provavelmente tiveram contatos intermitentes durante o Cretáceo. A Austrália por sua vez esteve em contato com a América do Sul durante praticamente todo o Cretáceo.
A evolução biogeográfica dos Marsupiais não asiáticos, parece ter começado no norte dos Andes onde são hoje encontrados os Marsupiais mais conservadores do planeta, ou seja. os Caenolestidae (representado por Caenolestes fuliginosus, a espécie mais conservadora). Como já demonstrei nas pranchas que publiquei neste álbum do FLICKR, esses marsupiais ainda conservam uma estrutura plesiomórfica no calcanhar muito semelhante àquela de Sinodelphys e muito mais conservadora que qualquer outro Marsupial vivo ou fóssil fora da Ásia.
Os Marsupiais Australianos e Sul-americanos (exceto Caenolestidae) compartilham um tipo de calcanhar derivado apresentando um astrágalo mais triangular que pode ser visto nesta figura e comparado ao dos Caenolestidae. Dentre os Caenolestidae conhecidos, Caenolestes fuliginosus pode ser considerado o mais conservador de todos eles. Sua distribuição geográfica é no norte dos Andes, aproximadamente entre a Baia de Guayaquil e o extremo norte da cordilheira. Exatamente a região antiga dos Andes. Também outra interessantíssima distribuição é a de Dromiciops gliroides exatamente na região por mim denominada Andes II. Dromiciops como já vimos é o grupo irmão dos Didelphimorphia conservando algumas características plesiomórficas já modificadas nos demais, como por exemplo o não encavalamento do incisivo 3 inferior (i3 stagged, na figura, uma sinapomorfia importante dos Didelphimorphia).
Veja "Hershkovitz, P. 1992a. Ankle bones: The Chilean opossum Dromiciops gliroides Thomas, and marsupial phylogeny. Bonner Zool. Beiträge 43 (2): 181-213".
Veja "Hershkovitz, P. 1992a. Ankle bones: The Chilean opossum Dromiciops gliroides Thomas, and marsupial phylogeny. Bonner Zool. Beiträge 43 (2): 181-213".
Finalmente os Didelphimorphia, que representam os Marsupiais Sul-americanos (exceto Caenolestidae e Dromiciops) mais os Australianos, se originaram nessa nova formação dos Andes que chamei de Andes III, durante o Cretáceo. Os Marsupiais fósseis da América do Norte e Europa, considerando suas características referentes ao calcanhar, provavelmente se originaram a partir das populações do norte da América do Sul, pois não compartilham as sinapomorfias citadas para os Didelphimorphia. Hershkovitz no entanto apontou um encavalamento do incisivo 3 em alguns fósseis do hemisfério norte, o que significa que pelo menos parte dos marsupiais que colonizaram a América do Norte eram mais aparentados com os Didelphimorfia. Hershkovitz também incluiu os Caenolestidae entre os Didelphimorphia no entanto o grande desenvolvimento do i2 nestes Marsupiais deixa essa conclusão parecer duvidosa.
This figure also explains the distribution of the genus Araucaria (the Paraná Pine) and related species, and that of the genus Nothofagus, in the southern Andes, New Zealand, New Caledonia, Australia and New Guinea. Such distribution is not result of dispersion, there are many elements associated with those areas where there is Nothofagus, such as marsupials, Araucareaceae, etc, lists have been published in several papers of botany and biogeography. It is fragmented biomas that evolved when the southern continents were join together. The Ratites birds were part of this associated bioms as Nothofagus-Araucaria forests and southern steppes.
 |
| Foto de Didelphoidea: Marmosa : John Harold Castano. |
Dromiciops é um gênero conservador sob alguns aspectos. Ele é o único marsupial vivo que apresenta uma cloaca "basicaudal" que só é encontrada em répteis e nos Monotremata. Os incisivos inferiores (mandibulares) apresentam-se igualmente espaçados e não encavalados como no caso do i3 dos Dasyuroidea e Didelphoidea (Caenolestoidea não oferece uma boa comparação), Hershkovitz, P. 1992a. Ankle bones: The Chilean opossum Dromiciops gliroides Thomas, and marsupial phylogeny. Bonner Zool. Beiträge 43 (2): 181-213.
Distribuição de caracteres em mosaico

Certain distributions of characters, such as those found on the basis of the evolution of marsupials, are very intriguing.
"The evolution is expressed much better through an analysis along the axes of dispersal of species, than through cladograms."
Caenolestidae and Didelphidae have paired spermatozoids, "a priori" this unique feature by itself should resolve the relationships of these two groups. However after a parsimonious analysis, we are forced to opt for convergence.
Maybe if we stop to think this might be a case of "Distribution in Mosaic." But the analysis does not stop there. Dromiciops has a foot intermediary between Didelphidae and marsupials with syndactyly, like Diprotodontia. But in turn not having teeth i3 stagged, like Didelphoidea and Dasyuroidea. In Marmosa astragalus has three areas (facets) well defined in plantar view: Calcaneus-astragalar (CaA), Sustentacular (Su) and astragalus-navicular (AN). In Sinodelphys and Caenolestes (Su) and (AN) are continuous, and separate from (CaA). In Dromiciops and Dasyuroidea (Su) and (CaA) are united (continuous).
If we think that this apparent confusion is merely remnants of characteristics shared by contiguous populations that resulted in a mosaic distribution, perhaps we could be approaching an important understanding to definitely understand the whole evolution of Marsupials.
If we start from the north to the south, "Caenolestes fuliginosus" that represent the most conservative species of Caenolestidae, have paired spermatozoids and are geographically contiguous with Didelphidae, but not with Dromiciops. Dromiciops lives in the southwestern Andes and has merged astragalar facets as Dasyuroidea (that are from Australia, western region) and a strong tendency to syndactyly. Didelphoidea and Dasyuroidea originated in the central Andes and share i3 stagged. This suggests heritages from a ancient Mosaic Distribution.
In one extreme Dromiciops have a basicaudal cloaca that is present only in reptiles and in Monotremata, and incisive of mandible evenly spaced.
Probably 2 ancient characters preseved in a peripherical distribution.
In the other extreme Caenolestes fuliginosus the most coservative Caenolestidae (and also the most conservative living marsupial) having several ancient characters like the heel bones, small eyes, non prehensile tail, homogenous coloration in pelage and more terrestrial hands and feet.
THE CHARACTERS ARE THERE...IT IS THE EXACT DISTRIBUTION.....IT DESCRIBES THE BEHAVIOR OF THE CHARACTERS.....DESCRIBES the behavior of the NATURE ! LIKE IT IS !
Para construirmos uma FILOGENIA, (A) podemos começar com uma triagem básica "gradista". Separando por similaridade. (B) Num segundo momento organizamos por grupos monofiléticos (baseados em sinapomorfias). (C) Depois disso, temos que examinar a possibilidade de existirem distribuições em mosaico. Pois aquilo que parece discordante, pode ser exatamente o avanço no conhecimento. A obstinação em resolver as "dicotomias" é negativa, pois as tricotomias existem. Finalmente temos que ter em mente, que os "CLADOGRAMAS" são muito úteis para se apresentar ideias, mas nunca irão refletir a evolução. Os cladogramas são esquemas LINEARES que estão desde a sua origem ligados a ideia de tempo, enquanto a evolução biogeográfica é espacial. Quando tentamos sobrepor "cladogramas de tempo" com "cladogramas de área", forçando, socando, "cortando pedaços"...estamos distorcendo as conclusões !
Eventualmente, um cladograma pode até representar exatamente a evolução de um grupo. Mas isso irá confundir, pois nossa mente extrapola e isso não é didático. Essa representação acima foi a maneira que criei para retratar a evolução precoce dos Metatheria. Sem dúvida esse retrato é melhor que um cladograma !
Calcaneus Evolution in MARSUPIALS
 |
Na ilustração acima utilizei desenhos de Szalay.
Szalay, F. S. 1994. Evolutionary History of the Marsupials and an Analysis of Osteological Characters. New York: Cambridge University Press, 1995. 481p. |
A relação entre Glironia e Caluromys é indiscutível. A posição intermediária de Marmosa e a unidade dos grupos australianos também são muito claras.
The Caenolestes position is very clear in this figure of course it is the sister-group to all other marsupials. The relationship between Glironia and Caluromys is indisputable. The intermediate position of Marmosa and the unity of Australian groups are also very clear.
Calcaneus evolution in Didelphoidea
 |
Na ilustração acima utilizei desenhos de Szalay e Zhe-Xi Luo. Szalay, F. S. 1994. Evolutionary History of the Marsupials and an Analysis of Osteological Characters. New York: Cambridge University Press, 1995. 481p.
|
Para compreendermos as relações filogenéticas existentes entre as espécies de Monodelphis, é necessário estabelecermos um quadro das possíveis relações existentes entre os demais marsupiais e esse gênero. A necessidade de um grupo externo para a polarização dos caracteres pede uma organização, pelo menos parcial, das idéias existentes à cerca da evolução dos marsupiais.
Os Didelphidae [sensu lato] têm sido considerados por muitos autores como os mais conservativos marsupiais existentes, "the Didelphyidae, a family which is otherwise so completely prototypal..." (Bensley, 1903), "the smaller Didelphidae are struturally prototypal to the remaining Marsupials" (Gregory, 1910: 217), "generalized" (Simpson, 1935), "the didelphyds are in all essentials the most primitive of the know true marsupials " (Winge, 1941), "Didelphis, as an example of a "living fossil""(Clemens, 1968: 1),"being considered satisfactory progenitors for the rest of the marsupials" (Kirsch & Calaby, 1977), "many didelphines are conservative in aspects of their structure, and their association within the subfamily is too often based on presence of primitive characters (symplesiomophies) and not on shared derived characters (synapomorphies)" (Marshall, 1982).
Se a família destacou-se da base do tronco evolutivo dos Marsupialia e não pode, no momento, ser considerada como um grupo monofilético, o gênero Monodelphis pode ter se originado diretamente da forma ancestral dos Marsupialia. Sendo assim, uma questão muito importante deve ser levantada.
Seriam os marsupiais um grupo monofilético? Para tentar responder a essa questão, partirei para uma especulação a esse respeito.
EVOLUTION and CLASSIFICATION of the MARSUPIALIA

The PHYLOGENY of MARSUPIALS

Com base em 2 trabalhos reuni 14 caracteres que apoiam a monofilia dos Marsupialia. Marshall (1979: 400) propôs um cladograma que mostra as relações filogenéticas existentes entre Prototheria, Metatheria e Eutheria, propondo sete caracteres sinapomórficos que justificariam a monofilia dos Marsupialia [1-7], pelo menos dos atuais. Adiante são discutidos mais três [8-10]. Hershkovitz (1992: 206) propõe 3 novos caracteres [11-14] (caracteres em negrito [n]).
Lista dos caracteres basais dos MARSUPIALIA:
1 - Alisfenóide como elemento principal na formação da bula auditiva ossificada.
2 - Desenvolvimento de uma "pseudovagina".
3 - O esmalte dos dentes possui "túbulos".
4 - Entre os dentes pós-caninos, só os últimos pré-molares tem predecessores.
5 - Hipoconúlido e entocônido próximos.
6 - Cúspide estilar C (l) grande e bem desenvolvida ( l sensu Hershkowitz, 1977) .
7 - Perda da cúspide C na pós-metacrista.
8 - Hálux oponível.
9 - Perda da garra do hálux.
10- Sobreposição do astrágalo sobre o calcâneo, com perda do contato entre a fíbula e o calcâneo.
11- Symphysis menti arredondada.
12- Perda do i1.
13- Mandíbula encurtada, originando perda de i1.
14- Posição dos testículos e da bolsa escrotal, anterior ao pênis.
Quatro caracteres são discutidos por Marshall (1979: 400), mas não são utilizados, chamo a atenção para os de número 32 e 22.
O caráter (32) reporta-se a antiga questão referente aos hábitos arborícola ou terrestre do ancestral Theria. Na tabela 1 (Marshall, 1979: 398) os Metatheria são considerados como primitivamente arborícolas, porém no cladograma da página 400, esse caráter não é utilizado.
A idéia de um ancestral arborícola para os marsupiais atuais foi primeiramente proposta por Huxley (1880) e posteriormente suportada por Dollo (1899), Bensley (1903), Tate (1933), Lewis (1964) e mais recentemente por Szalay (1984). Szalay baseou-se principalmente num estudo detalhado das relações existentes entre os elementos tarsais, tíbia e fíbula de diversos mamíferos.
Entre os caracteres utilizados por vários desses autores destacam-se: o hálux oponível sem garra; a redução dos artelhos II e III, acompanhada de sindactilia e predominância do artelhos IV em muitas das espécies australianas e a completa sobreposição do astrágalo sobre o calcâneo acompanhada da perda do contato entre a fíbula e o calcâneo, com o aparecimento de um meniscus entre a fíbula e o astrágalo. Esses caracteres poderiam ser utilizados na análise cladística efetuada independentemente da discussão à cerca do hábito do ancestral Theria.
O hálux oponível, presente em muitos dos marsupiais atuais, não aparece nos Prototheria, nem nos grupos considerados mais precoces de Eutheria, podendo ser adotado como uma sinapormorfia dos marsupiais atuais. A ausência de garra no hálux merece uma discussão mais detalhada, pois os Notoryctidae apresentam uma garra no hálux (caso único entre os marsupiais). Essa condição a priori poderia ser considerada como plesiomórfica, por representar a condição do ancestral dos mamíferos. No entanto, Dollo (1899: 200) mostra que Notoryctes exibe caracteres que indicariam uma origem arborícola tais como: predominância do artelho IV (6), redução dos artelhos II e III, ligeira sindactilia nesses artelhos e entocuneiforme alongado, o que não seria de se esperar numa pata encurtada. As três primeiras características estariam numa condição derivada no contexto dos marsupiais atuais, e são compartilhadas exclusivamente pelos Syndactyla (Peramelidae e Diprotodontia) devendo ser considerada como uma sinapomorfia que agruparia Notoryctidae aos Syndactyla. Dollo (1899: 202) também faz comparações entre Peramelidae e Notoryctidae, citando alguns caracteres que poderíamos considerar como sinapomórficos para esses grupos: caixa timpânica volumosa, patela ossificada (5) e marsúpio voltado para trás (1). Ainda baseado nas informações fornecidas por Dollo sobre esses grupos, podemos acrescentar mais três sinapomorfias; a presença de um entocuneiforme alongado, paralelo ao eixo maior da pata (2); redução do mesocuneiforme (4) e um contato entre o metatarso V e o calcâneo (3). Essas evidências indicam que esses dois grupos apresentam estreitas relações de parentesco, formando um grupo monofilético. Esses dois grupos formariam um táxon, que juntamente com seu grupo irmão, os Diprotodontia formariam os Syndactila. (fig. 71).
Szalay (1984: 255) afirma que o arranjo cruro-tarsal e intertarsal da pata de Notoryctes, retêm o mesmo padrão encontrado nos demais marsupiais australianos, padrão esse que ele considera derivado da condição ancestral dos marsupiais atuais. Com base nesse padrão e na perda do incisivo 4 inferior podemos considerar que todos os marsupiais australianos formariam um grupo monofilético, englobando finalmente os Dasyuridae.
Essas evidências corroboram a idéia de que Notoryctidae não representa um ramo (precoce) basal na filogenia dos marsupiais atuais. Dessa maneira a garra presente em Notoryctidae pode ser, por uma questão de parcimônia, interpretada como uma autapomorfia desse grupo e não como uma condição ancestral retida. A ausência de garra nos marsupiais [perda], pode ser considerada como uma sinapomorfia destes, e a presença de unhas nos artelhos de Notoryctidae, seriam perfeitamente explicadas pela drástica paedomophosis, que marcou sua evolução.
A completa sobreposição do astrágalo sobre o calcâneo, a perda do contato entre a fíbula e o calcâneo com o aparecimento de um meniscus entre a fíbula e o astrágalo, representam condições plesiomórficas para os marsupiais australianos e para Dromiciops (Szalay, 1984: 235-236).
De acordo com os argumentos apresentados por esse autor, uma série de transformações pode ser aqui utilizada para estabelecer sinapomorfias em dois níveis de universalidade diferentes. A condição encontrada nos "Didelphidae" [condição ancestral] pode ser considerada uma sinapomorfia para todos os marsupiais atuais e representa uma evidência a favor da idéia relacionada aos hábitos arborícolas desse ancestral. A sindactilia observada em várias espécies australianas deve ser utilizada apenas para se definir Syndactyla.
Voltando ao ponto inicial da discussão do caráter (32) de Marshall (1979: 393), podemos dizer que independentemente da decisão que tenha sido adotada por diferentes autores quanto aos hábitos arborícolas ou terrestres do ancestral Theria, parece pouco provável que o ancestral dos marsupiais atuais não tenha sido arborícola, já que a anatomia aponta fortemente nessa direção.
O caráter (22) de Marshall (1979: 382) refere-se a posição dos testículos e a bolsa escrotal. Podemos nos valer da mesma discussão já apresentada quando tratei da presença de uma garra no hálux de Notoryctes. Se todos os marsupiais atuais apresentam a bolsa escrotal anterior ao pênis, condição esta que não está presente em Notoryctes, no qual os testículos possuem posição intra-abdominal, a condição encontrada nesse gênero, por parcimônia, deve ser considerada como autopomórfica, pois seis caracteres diferentes corroboram a hipótese que Notoryctes não representa um ramo precoce (basal) na cladogênese dos marsupiais.
Dessa discussão podemos retirar três sinapomorfias para marsupiais. As quais juntamente com aquelas propostas por Marshall (op. cit.) vêm corroborar a hipótese sobre a monofilia ao menos dos marsupiais atuais: hálux oponível [8], perda da garra do hálux [9] e sobreposição do astrágalo sobre o calcâneo, com perda do contato entre a fíbula e o calcâneo [10] (fig. 72).
Uma vez estabelecida a monofilia dos marsupiais, resta-nos examinar os agrupamentos internos. Por uma questão de conveniência tratarei primeiro dos marsupiais australianos.
Como já foi visto na análise feita por Szalay (1984). As patas posteriores [pes] dos marsupiais australianos, assim como a de Dromiciops, exibem um padrão derivado da condição encontrada nos Didelphidae [sensu lato]. O contato entre o astrágalo e o calcâneo está modificado. O sulcus calcanei está ausente pois as facetas calcâneo-astragalar e sustentacular (faceta calcâneo-astragalar distal) estão fundidas. Tal condição representa uma sinapomorfia dos Dasyuroidea [=Australidelphia] [81]. Os Dasyuroidea [=Australidelphia] não apresentam mais que seis incisivos inferiores, ou seja, os incisivos inferiores podem estar reduzidos em graus variados mas nunca ultrapassam seis. A presença de oito incisivos inferiores, como nos "Didelphidae", é a condição ancestral dos marsupiais, Thomas (1888a: 315), Winge (1841: 101) e mais recentemente (Woodward, 1893; Osgood, 1921; Takahashi, 1974) (apud Hershkovitz, 1982). Considerada como plesiomórfica para os Marsupialia (Marshall, 1979: 388). A perda do I4 nos Dasyuroidea pode ser considerada uma sinapomorfia destes [82]. Portanto duas sinapomorfias podem ser utilizadas para justificar a monofilia dos Australidelphia (fig. 72).
Dentro de Dasyuroidea podemos reconhecer 2 grupos monofiléticos os Syndactyla [Diprotodontia + Perameloidea], com sindactilia [84] e os Dasyuridae, com redução progressiva do artelho I. [83]
Diprotodontia é seguramente monofilético incluindo "Phalangeroidae" [parafilético] + Macropodidae [monofilético se incluírmos os Potoridae]. Caracteres: diprotodontia e diapausa embrionária [85-86]
Perameloidea incluindo Notoryctidae [monofilético] + Peramelidae [parafilético, justamente por não incluir os Notoryctidae]. Caracteres na página 135, fig 71. [87-92] na fig 72.
Mirmecobiidae e Thylacynidae devem ser apenas membros mais diferenciados de Dasyuridae, que foram separados por grau de evolução, deixando para trás um ramo parafilético. Os Dasyuridae mostram claramente uma evolução das patas posteriores com redução progressiva do artelho I, distanciando-se o padrão ancestral.
Hershkovitz 1992 forneceu mais caracteres que corroboram a monofilia dos marsupias além discutir a posição filogenética de Dromiciops e sua relação com os marsupiais australianos (fig. 72).
As evidências apresentadas por Hershkovitz (1992 : ) a respeito da posição ocupada por Dromiciops na filogenia dos marsupiais são sem dúvida alguma as mais bem fundamentadas até hoje. Em minha filogenia de 1991 inclui Dromiciops nos Marmosidae com base nas características cranianos justificaveis. Mas agora diante das novas evidências devo reconhecer que as características que usei devem ser consideradas como convergências, devidas provavelmente a uma evolução pedomórfica. Pois de um modo geral todos os filhotes dos Didelphidae [sensu lato] apresentam caixa craniana arredondada, focinho curto órbita grande e os foramens do basicrânio muito próximos, características estas que se conservam nos adultos dos Marmosidae.
Certamente o ponto mais polêmico dessa filogenia é a posição de Dromiciops. Este gênero possui características derivadas compartilhadas com os marsupiais australianos relacionadas à evolução do pé (vários aspectos) e plesiomórficas como o tipo de espermatozoide.
No entanto apresenta certas características conservadoras como uma cloaca basicaudal (como os Monotremata e os répteis), incisivos da mandíbula não encavalados (que estão modificados nos Dasyuroidea e nos Didelphoidea), e outras levantadas por Herkovitz in: "Hershkovitz, P. 1992a. Ankle bones: The Chilean opossum Dromiciops gliroides Thomas, and marsupial phylogeny. Bonner Zool. Beiträge 43 (2): 181-213".
Estas características plesiomórficas colocariam Dromiciops antes dos Dasyuroidea e dos Didelphoidea, já que esses 2 grupos compartilham várias características derivadas.
Echymipera e Notoryctes formam um grupo monofilético separado de Peramelinae, que está sendo temporáriamente considerado como Notoryctidae "PARA CHAMAR A ATENÇÃO".
DASYURIDAE EVOLUTION based on calcaneus-astragalus facets

Here phylogenetics relationships of some Dasyuridae, based on Szalay studies of calcaneus-astragalus facets. I tentatively reunite Myrmecobius and Thylacinus, but I think it's problematic. I tentatively reunite Myrmecobius and Thylacinus, but I think it's problematic. Szalay considered as dasyuroids what I'm considering as Dasyurids, cited in the plate.

Myrmecobius e Thylacinus não se assemelham apenas na belíssima pelagem, mas também nas características dos elementos tarsais do pé.

Aparentemente Echymipera é parafilético, mas o que é mais estranho é que
Echymipera doryanus é considerada subspécie de Echymipera kalubu [doreyanus].
Echymipera doryanus é considerada subspécie de Echymipera kalubu [doreyanus].
Evolution of the Foot in Marsupials

Na ilustração acima utilizei desenhos de Szalay .
Szalay, F. S. 1994. Evolutionary History of the Marsupials and an Analysis of Osteological Characters. New York: Cambridge University Press, 1995. 481p.

Nesse quadro coloquei Notoryctes dentro da família Echymiperinae pelas afinidades filogenéticas existentes entre esse gênero e Echymipera. Pois um animal como a toupeira marsupial deve ter vindo de um animal escavador.
Sempre posicionam animais diferentes como se viessem do NADA ! Separados !
Sabemos que animais muito "diferentes" só podem vir de animais comuns, é assim que funciona.
O padrão básico para todos os marsupiais Australianos é um animal parecido com Marmosa. A partir daí surgiram 3 linhas:
1- Dasyuridae : insetívoros que evoluíram também para carnívoros (pata adaptada para uma vida no chão) têm o calcânio derivado e redução do artelho 1.
2- Peramelidae : desceram das árvores (pois têm sindactilia) tornaram-se escavadores...daí vem seu alimento e tocas (de proteção e cria). Evoluíram para "toupeiras".
3- Diprotodontia : primitivamente arborícolas com dois dentes longos na frente da mandíbula que existem em outros pequenos insetívoros de diversos grupos, inclusive nos Rodentia. Se tornaram os maiores herbívoros da Austrália.
NOTA : Se alguém quiser colocar os Caenolestidae "dentro" dos Diprotodontia...não confie nessa pessoa como zoólogo ! Logo logo estará colocando roedores também !
In this picture I put Notoryctes within the family Echymiperinae by the phylogenetic affinities with this genre and Echymipera. Think about it !
An animal like a marsupial mole must have come from a digger animal.
Always different animals are positioned as if they came from NOTHING! Separated!
We know that animals much "different" can only have come from normal animals, it is so !
The basic pattern for all Australian marsupial is an animal resembling Marmosa.
From this emerged three lines:
1 - Dasyuridae: insectivores that have also evolved for carnivores (foot adapted for a life on the ground) they have a calcanium derived and toe 1 reduced.
2 - Peramelidae: they came down from the trees (because they have syndactyly) became diggers ... hence their food and burrows (protection and offspring). Evolved into "moles".
3 - Diprotodontia: primitively arboreal with two long incisives at the tip of the jaw
they are also found in other small insectivores of several groups, including the Rodentia. Became the largest herbivores in Australia.
NOTE: If someone wants to put Caenolestidae "inside" Diprotodontia ... do not trust this person as a zoologist! Pretty soon will be putting rodents too!
Calcaneus-astragalus evolution in DIPROTODONTIA

Compreender a função de cada uma das partes do calcâneo e do astrágalo é fundamental para perceber a maneira como os marsupiais evoluíram. Uma das razões que motivou o grandioso trabalho de Szalay foi o fato desses ossos se preservarem bem como fósseis. Através desse conhecimento comparativo pode-se traçar toda a linha evolutiva dos marsupiais desde a sua origem.
Esse estudo se perpetua no trabalho de Zhe-Xi Luo, seus colegas e colaboradores.
Vejam a simplificação [pedomórfica] do astrágalo em Tarsipes, resultou numa forma próxima a do ancestral, esse tipo de regressão pode confundir em uma análise (em algumas etapas iniciais pelo menos).
Todas as evidências mostram que Tarsipes sofreu uma redução de tamanho, levando a uma série de autapomorfias, o calcaneus simplesmente se simplificou.
Além da aparência externa e da pelagem, Marmosa também se mostra conservadora na forma do calcaneus e Eudromicia é o mais conservador dos Dasyuroidea, também nesses aspectos.
Dromiciops mostra uma convergência muito grande com Burramys no calcaneus, porém uma reunião destes gêneros não se sustentaria obviamente. Isso sugere uma condição derivada em Dromiciops não só com relação a fusão das facetas calcâneo-astragalares, mas também no contato com o cubóide (cor violeta).
A relação calcâneo-astrágalo mostra-se mais conservadora nos Diprotodontia basais que nos Dasyuridae ou nos Peramelidae (lato sensu), por isso escolhi essa comparação, esses dois grupos se distanciaram mais do ancestral arborícola.
Na ilustração acima utilizei desenhos de Szalay e Zhe-Xi Luo.
Szalay, F. S. 1994. Evolutionary History of the Marsupials and an Analysis of Osteological Characters. New York: Cambridge University Press, 1995. 481p.
Understanding the role of each of the parts of the calcaneus and the talus is essential to understand how the marsupials evolved. One of the reasons that motivated the great work of Szalay was the fact that these bones preserve well as fossils. Through this comparative knowledge can trace the entire evolutionary line of marsupials since its origin.
This study is perpetuated in the work of Zhe-Xi Luo, colleagues and collaborators.
See how the simplification [pedomorphosis] in Astragalus of Tarsipes resulted in a form close to the ancestor, this type of regression can be confusing in a analysis (in some stages at least).
All the evidence suggests that Tarsipes size was reduced, leading to a series of autapomorphies, the calcaneus simply simplified.
In addition to the external appearance and coat, Marmosa also shown the conservative form of the calcaneus and Eudromicia is the most conservative Dasyuroidea, also in these respects.
Dromiciops shows a great convergence with Burramys in the calcaneus, but a reunion of these genres obviously is not sustainable. This suggests a derived condition in Dromiciops not only with the fusion of the calcaneus-astragalus facets, but also in contact with the cuboid (violet).
The relationship calcaneus-astragalus appears to be more conservative in the baseline Diprotodontia, than in Dasyuridae or Peramelidae (sensu lato), so I chose this comparison, these two groups diverged more from the arboreal ancestor.
The Evolution of Diprotodontia

Para começar vamos falar dos Diprotodontes por ser o grupo mais diversificado de todos.Nesse grupo estão os cangurus que representam no imaginário popular o modelo de marsupial. Estão também os Coalas famosos pela aparência de urso de pelúcia.
Cohort: Didelphimorphia - i 5/4, c 1/1, p 3/3, m 4/4 = total 50
-Dasyuroidea - i 4/3, c 1/1, p 3/3, m 4/4 = total 46
-Diprotodontia - i 3/2, c 1/0, p 3/3, m 4/4 = total 40
Os diprotodontes perderam incisivos e os caninos inferiores. Ao longo de sua evolução perderam dentes e estes tornaram-se mais especializados de acordo com o tipo de alimento utilizado.
Na figura acima acompanhe as perdas de dentes [em vermelho].
O coala Phascolarctos [Phascolarctidae] voltou para as árvores partindo de um ancestral terrestre mas manteve caracteres que atestam essa passagem : marsúpio voltado para trás, olhos pequenos, focinho rombudo e redução da cauda, como em seu grupo irmão Vombatidae.
Cohort: Didelphimorphia: 1- Dasyuroidea, 2- Diprotodontia
Na figura acima acompanhe as perdas de dentes [em vermelho].
O coala Phascolarctos [Phascolarctidae] voltou para as árvores partindo de um ancestral terrestre mas manteve caracteres que atestam essa passagem : marsúpio voltado para trás, olhos pequenos, focinho rombudo e redução da cauda, como em seu grupo irmão Vombatidae.
Cohort: Didelphimorphia: 1- Dasyuroidea, 2- Diprotodontia
The Evolution of Diprotodontia (skulls) part 1

The skulls from the site of Museum Victoria:

A familia Burramyidae assim como o gênero Cercartetus não representam um grupo monofilético.
O gênero Eudromicia é basal para todos os Diprotodontia, sem dúvida alguma seu crânio com focinho alongado contrasta com os focinhos encurtados de todos os gêneros basais como Burramys, Acrobates, Petaurus e Cercartetus onde costuma ser incluído . Também com base no aspecto externo e padrão da pelagem pode ser considerado como o mais conservador de todos os marsupiais da Australásia. Sua semelhança com o gênero Marmosa, que também é basal para os Didelphoidea, e com Dromiciops não nos deixa melhor alternativa. Suas modificações em relação ao padrão ancestral são também as sinapomorfias dos Diprotodontia : sindactilia e incisivos diprotodontes.
Dromicia concinna por sua vez está na base dos gêneros de tamanho médio como Petaurus e Gymnobelideus e como eles possui o arco zigomático inflado e oco na região da fossa glenóide, essa característica representa uma modificação chave na filogenia de todos os demais diprotodontia.
Resta então na família Burramyidae apenas 3 espécies: Burramys parvus, Cercartetus nanus e C. lepidus.
As famílias Acrobatidae e Tarsipedidae devem ser consideradas como grupos irmãos unidos com base na redução do processo coronóide da mandíbula assim como o estreitamento do ramo horizontal que em Tarsipes se reduziu a um bastão.
Essas duas famílias mais os Burramyidae formam um grupo monofilético com adaptações para uma dieta especializada em pólem e néctar, com línguas modificadas providas de papilas longas formando uma estrutura em forma de escova. Uma redução no tamanho do corpo também permite que explorem ramos mais delicados onde ficam as flores. Burramys tem tamanho maior e alimentação derivada com dentes especializados.
Tarsipes é um dos gêneros mais modificados entre os Diprotodontia, por isso a mentalidade "gradista" o colocou num grupo separado e a análise molecular está fazendo o mesmo, por separar tudo o que diverge em termos de distância genética.
Mas como distância genética não é sinônimo de distância filogenética, acabamos no retrocesso "gradista".
The family Burramyidae or the genus Cercartetus do not represents a monophyletic group.
The genus Eudromicia is basal to all Diprotodontia undoubtedly his skull with elongated snout contrasts with shortened muzzles of all basal genres as Burramys, Acrobates, Petaurus and Cercartetus in which is usually included. Also based on the pattern and external appearance of the fur can be considered as the most conservative of all australasian marsupials. His resemblance to the genus Marmosa, which is also the baseline for Didelphoidea, and Dromiciops, does not leave us best alternative. Their modifications from the ancestor are also Diprotodontia synapomorphies: syndactyly and diprotodont incisors.
Dromicia concinna in turn is at the base on the medium size genera as Petaurus and Gymnobelideus, and equaly have a hollow zygomatic arch inflated in the region of the glenoid fossa, this feature is a key modification in the phylogeny of all Diprotodontia.
There remains in the family Burramyidae only 3 species: Burramys parvus, Cercartetus nanus and C. lepidus.
Families Acrobatidae and Tarsipedidae should be considered together sisters groups based on the reduction of the coronoid process of mandible and the narrowing in the horizontal ramus, that is in Tarsipes reduced to a stick.
These two families plus Burramyidae form a monophyletic group with adaptations for a specialized diet on pollen and nectar, with modified language provided with long papillae forming a brush-shaped structure. A reduction in body size also allows them to explore more delicate branches where the flowers are. Burramys have larger size and derivative feeding with specialized teeth.
Tarsipes is one of the most modified genera among Diprotodontia, so the “gradist” mentality put him in a separate group and molecular analysis is doing the same, by separating all that differs in terms of genetic distance.
But how genetic distance is not synonymous of phylogenetic distance, we just step in a "gradistic" backwards.
The Evolution of Diprotodontia (skulls) part 2

Nesse quadro estão os marsupiais mais famosos do planeta, O CANGURU e O COALA !
O grupo irmão deles é a familia Phalangeridae, são marsupiais diprotodontes que vivem em árvores mas também vivem no chão. Foi justamente desse grupo marsupiais que evoluíram os Vombatiformes e os Macropodiformes.
Na base desse cladograma vemos um crânio de Trichosurus (Phalangeridae), a seguir uma dicotomia, à esquerda os Vombatiformes e à direita os Macropodiformes.
Phascolarctidae, Vombatidae, Hypsiprymnodontidae, Potoridae e Macropodidae: formam um grupo monofilético.
Esses animais representam como um todo, um estágio avançado da evolução dos marsupiais australianos, seu grupo irmão são os Phalangeridae (Trichosurus, Phalanger e Wyulda), outro grupo monofilético.
O coala é o único gênero da familia Phascolarctidae, e sua evolução foi uma das mais estranhas entre os marsupiais australianos. Esse animal compartilha vários caracteres derivados com os Vombatidae, seu grupo irmão mais imediato. Os vombates são escavadores e possuem dentes de crescimento contínuo como os roedores. Vive em tocas no subsolo.
Derivado de opossuns arborícolas com cauda preênsil, o COALA praticamente não tem cauda, pois sua evolução foi inicialmente terrestre e semi-fossorial, "desceu das árvores" depois voltou, mas nessa volta trouxe consigo as marcas de sua evolução com animal terrestre, que vivia em tocas no chão, olhos pequenos, ausência de cauda e uma marsúpio voltado para trás, como um marsupial escavador.
- Infelizmente a figura que consegui de Hypsiprymnodon moschatus, não é muito boa...peço a quem tiver uma melhor me enviar para melhorar a prancha ! Não é possível reunir todas as espécies numa só figura, devido ao "balanço", "equilíbrio" entre espaço e a didática. No entanto aqui podemos ver claramente as afinidades dos principais gêneros e como se deu a evolução desse grupo. A partir daqui fica mais fácil a compreensão dos gêneros restantes. Claro que me faltam também fotografias de vários gêneros, assim como informações sobre os ossos do pé.
- Infelizmente a figura que consegui de Hypsiprymnodon moschatus, não é muito boa...peço a quem tiver uma melhor me enviar para melhorar a prancha ! Não é possível reunir todas as espécies numa só figura, devido ao "balanço", "equilíbrio" entre espaço e a didática. No entanto aqui podemos ver claramente as afinidades dos principais gêneros e como se deu a evolução desse grupo. A partir daqui fica mais fácil a compreensão dos gêneros restantes. Claro que me faltam também fotografias de vários gêneros, assim como informações sobre os ossos do pé.
.... unfortunately the figure I got of Hypsiprymnodon moschatus, is not very good ... I ask whoever has a better send me to improve the FIGURE!

A hollow zygomatic arch is certainly an adaptation to arboreal life.
The smaller Burramyidae and Tarsipedide jump from flower to flower. The Acrobatidae jumpers became small gliders, with a line of hairs expanding the tail sideways.In Petauridae the zygomatic arch is robust but with a porous structure. Won resistance but not increased the body weight of these medium-sized arboreal gliders. A robustness in the glenoid region of the zygomatic arch is observed in all Diprotodontia from Petauridae forward (see the pictures related)
The skulls from the site of Museum Victoria:
Essa figura mostra a evolução do crânio dos Diprotodontes basais.
Um destaque para a redução da mandíbula que ocorreu em Tarsipedidae e Acrobatidae, uma adaptação a dieta de pólem e néctar.
Outro destaque para uma das principais modificações do crânio dos Diprotodotia é o desenvolvimento da região glenóide do arco zigomático. Essa área responsável pela inserção da musculatura ligada à mastigação é bem desenvolvida nas formas de maior tamanho. Nos Petauridae e Pseudocheiridae é possível visualizar a consistência porosa dessa área, provavelmente possibilitada uma leveza com resistência, fator importante para um animal arborícola principalmente os planadores e saltadores.
Dromicia concinna in turn is at the base on the medium size genera as Petaurus and Gymnobelideus, and equaly have a hollowed zygomatic arch inflated in the region of the glenoid fossa, this feature is a key modification in the phylogeny of all Diprotodontia.
There remains in the family Burramyidae only 3 species: Burramys parvus, Cercartetus nanus and C. lepidus.
Families Acrobatidae and Tarsipedidae should be considered together sisters groups based on the reduction of the coronoid process of mandible and the narrowing in the horizontal ramus, that is in Tarsipes reduced to a stick.

O quadro acima tenta mostrar a inconsistência de Burramyidae como tem sido apresentada até então e sua relação com Acrobatidae e Tarsipedidae. Notem como Eudromicia destacou-se antes do encurtamento do focinho. Dromicia concinna apresenta um arco zigomático inflado na região glenóide. Acrobates está numa posição intermediária entre Eudromicia e Tarsipes, tanto em relação à mandíbula como na forma do calcâneo e do astrágalo. Tarsipes representa um extremo de redução na mandíbula, é possível ver-se um esboço do processo coronóide e um resquício da curva do côndilo. Na ponta do ramus horizontal um incisivo. Por isso estou restringindo os Burramyidae a 3 espécies somente: Burramys parvus, Cercartetus nanus e C. lepidus.
The table above attempts to show the inconsistency of Burramyidae as has been shown so far and its relationship with Acrobatidae and Tarsipedidae. Notice how Eudromicia stood out before the shortening of the muzzle. Dromicia concinna has an inflated zygomatic arch in the glenoid region. Acrobates is an intermediate position between Eudromicia and Tarsipes, both in relation to the mandible and the manner of the calcaneus and talus. Tarsipes presents an extreme reduction in the jaw, it is possible to see an outline of the coronoid process and a remnant of the curve of the condyle. At the tip of horizontal ramus an incisor. So I'm restricting Burramyidae the three species only: Burramys parvus, Cercartetus nanus and C. lepidus.
Na ilustração acima utilizei desenhos de Szalay.
Szalay, F. S. 1994. Evolutionary History of the Marsupials and an Analysis of Osteological Characters. New York: Cambridge University Press, 1995. 481p.
The skulls from the site of Museum Victoria: https://collections.museumsvictoria.com.au/specimens/139273
Evolução dos dentes e da Alimentação nos DIPROTODONTIA
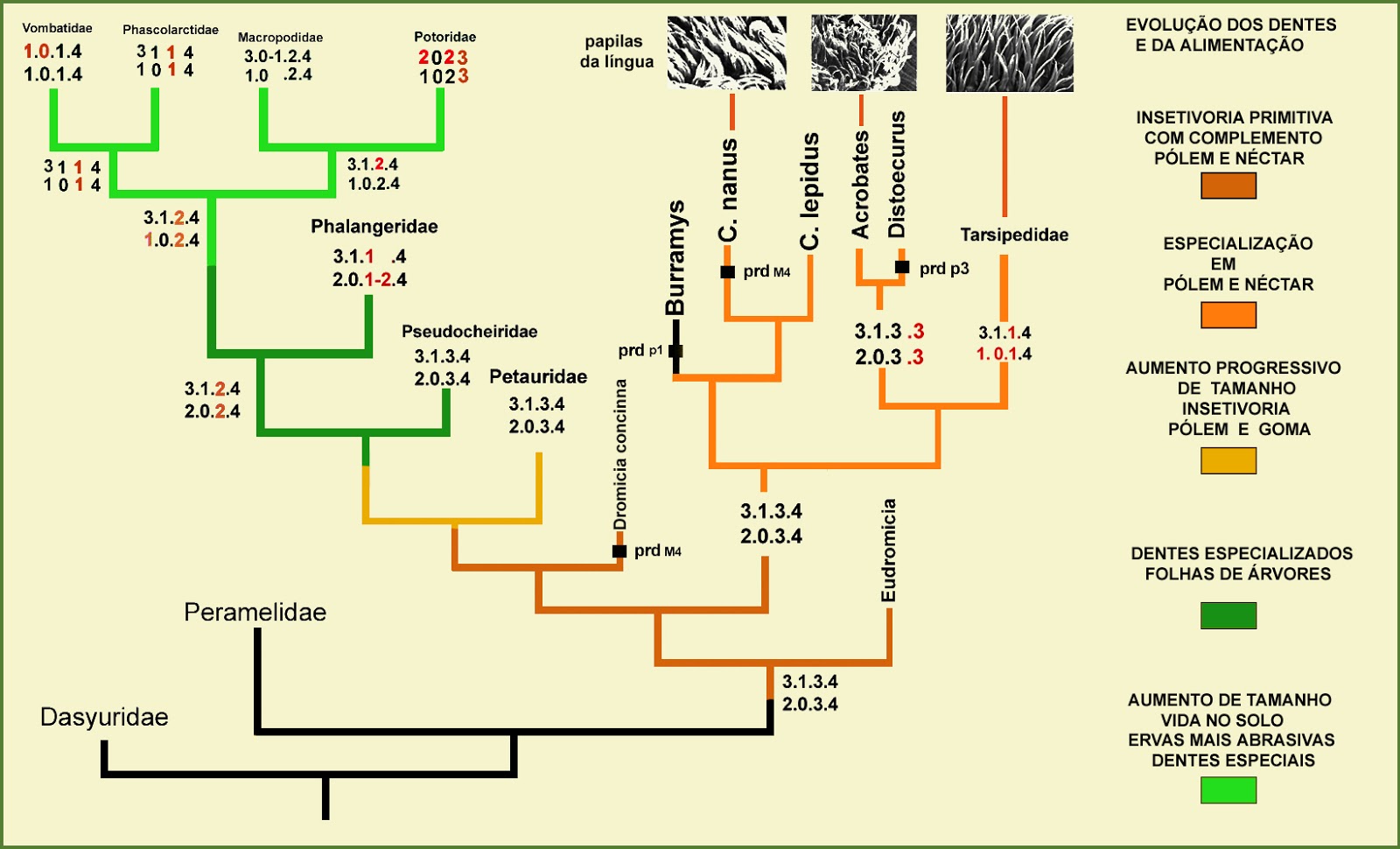
Fórmulas dentárias dos Marsupiais - incisivos. caninos. pré-molares. molares.
Cohorte: Didelphimorphia - i 5/4, c 1/1, p 3/3, m 4/4 = total 50
Dasyuroidea - i 4/3, c 1/1, p 3/3, m 4/4 = total 46
Diprotodontia - i 3/2, c 1/0, p 3/3, m 4/4 = total 40
Os diprotodontes perderam incisivos e os caninos inferiores. Ao longo de sua evolução perderam dentes e os restantes tornaram-se mais especializados de acordo com o tipo de alimento utilizado.
Na figura acima acompanhe as perdas de dentes [em vermelho].
O coala Phascolarctos [Phascolarctidae] voltou para as árvores partindo de um ancestral terrestre mas manteve caracteres que atestam essa passagem : marsúpio voltado para trás, olhos pequenos, focinho rombudo e redução da cauda, como em seu grupo irmão Vombatidae.
THE EVOLUTION of the Marsupial Digestive System
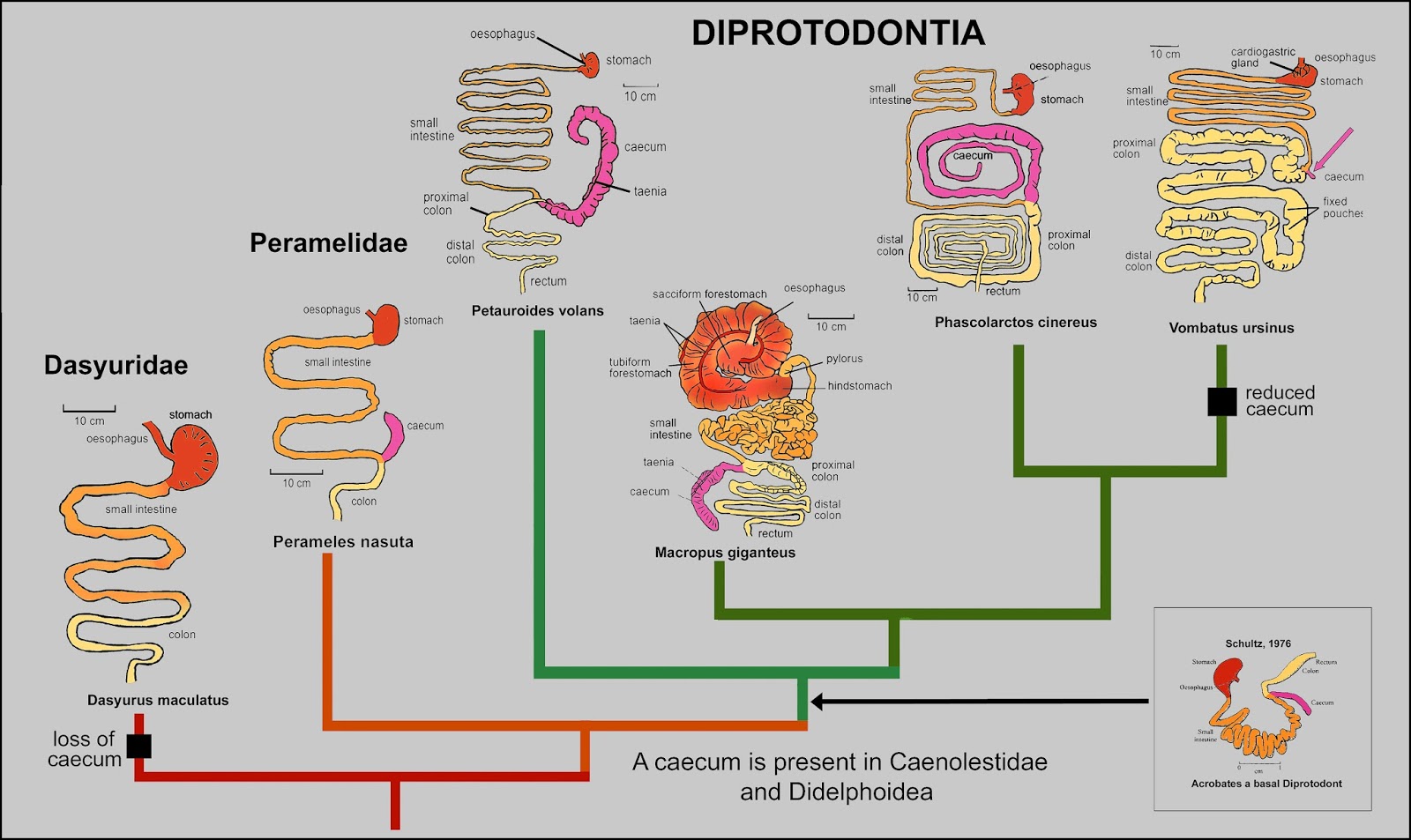
Modified from T.J. DAWSON, et al.- FAUNA of AUSTRALIA - MORPHOLOGY AND PHYSIOLOGY OF THE METATHERIA.
O sistema digestivo dos Dasyuridae se simplificou perdendo o caecum, adaptados a uma dieta insetívora e depois carnívora. O exemplo acima mostra o trato digestivo de Dasyurus maculatus que é praticamente igual a todos os membros da familia.
Embaixo à direita o trato digestivo de Acrobates, um diprotodonte basal, muito semelhante ao dos Peramelidae. Ao que tudo indica semelhante ao do ancestral de todos os marsupiais atuais, um onívoro com um pequeno caecum ligado ao intestino, que ajuda na digestão de substâncias que necessitam de ação bacteriana.
O trato digestivo se diversificou bastante dentro dos Diprotodontes, a partir de Petauridae com a introdução da goma na dieta provavelmente foram criadas as condições para uma evolução totalmente vegetariana.
Male genital system evolution in Marsupials

"Male echidnas ejaculate with just two heads (half of the penis) at a time. This resembles very much the way lizards and snakes ejaculate: they have a double penis (hemipenis), but only one of the two penises is used during the copulation, while the other will effectuate the next copulation/ejaculation. Marsupials are now in "this matter something between monotremes and placental mammals": they do not use half of the penis for mating, but still have a double headed penis, while the echidnas have a reptilian joined hemipenis, with each part of the penis in a marsupial-like fashion." Stefan Anitei in "Softpedia".
Nesse caso o penis do Echidna representa uma junção dos 2 hemipenises reptilianos. Isso explicaria o uso alternado das cabeças, exatamente como no caso do hemipenis dos répteis de uso alternado."Esse desenho do pênis de um Ornithorhynchus mostra uma assimetria, como se fosse meio pênis de Echidna. Possui papilas e uma uretra fechada diferente do que ocorre nos marsupiais, que possuem um sulco uretral reptiliano".
A bolsa escrotal dos marsupiais evoluiu independentemente dos Placentários. Nos marsupiais o escroto é pré-peniano e não pós-penianos como em muitos placentários. Elefantes, musaranhos e Monotremados possuem testículos intra-abdominais.
"The theory of the allometric fractals explains much about the variations and the way by which the evolution works".
EVOLUTION is a EMBRYONIC PROCESS ! a cut in the embryological process......or a prolongation in this process !
Didelphis e Philander apresentam característica penianas, que poderiam ser consideradas como basais, se comparadas às dos Marsupiais australianos. Nos australianos observamos uma redução gradativa da ponta bífida da glande. Nos Macropodidae existe uma simplificação máxima. Coalas e vombates apresentam uma situação intermediária, mais próxima dos Macropodidae. Em Caluromys, Micoureus e Marmosa observamos uma modificação da glande, que se apresenta alongada, provavelmente uma adaptação à vida arborícola.Os penises de Caluromys e Micoureus possuem a abertura externa da uretra localizada bem na ponta do órgão. Estes gêneros também possuem glândulas produtoras de mucos bem desenvolvidas na abertura da cloaca. Eu considero essas características como sinapomorfias de Caluromys e Micoureus, uma adaptação à vida arbórea. A cópula desses animais ocorre num ambiente onde as opções de posicionamento são bastantes limitadas. Por isso um penis mais alongado com abertura na ponta e muita muita mucosidade ajudam na transferência dos gametas. Em Marmosa temos uma situação um pouco mais conservadora com relação à abertura da uretra.
Outras evidências reforçam as estreitas relações filogenéticas entre Micoureus, Caluromys e Glironia: In most marsupial species there are "three pairs" of Cowper's (bulbourethral) glands, in Caluromys and Glironia there are "two pairs".
"The cloaca of Caluromys is the widest amongst South American marsupials studied." " The presence of a well-developed glandular complex around the cloacal cleft of Micoureus demerarae, Caluromys lanatus and Caluromys philander must be physiological significance in the processes of penile erection and exposure by lubrificating the wall opening to facilitate the sliding of the glans." Nogueira and Castro (2004) Predators with Pouches: The Biology of Carnivorous Marsupials: cap.2.
Análise dos elementos tarsais de Dasyuridae e Peramelidae

Quais são os pré-requisitos necessários para se analisar os elementos tarsais dos Dasyuroidea ?
Sinodelphys é importante não apenas por sua antiguidade, mas por sua posição basal dentro da evolução dos marsupiais. A antiguidade é uma questão de datação e está subordinada a isso. A sua posição basal foi reconhecida por comparação com Eomaia e outras formas semelhantes tanto na linha dos marsupiais como na linha dos Eutheria.
Caenolestes é importante por estar tanto morfologicamente como geograficamente a meio caminho entre Sinodelphys e os Microbiotheridae e Didelphimorfia, que juntos formam o grande grupo dos marsupias atuais com caudas preênsis, hálux sem unha e elementos tarsais mais avançados.
Tanto Sinodelphys como Caenolestes possuem duas facetas calcâneo-astragalares (em lilás). Nesse sentido os Didelphoidea são os mais conservadores e portanto servem como elementos de comparação, e entre eles sem dúvida alguma o gênero Marmosa é o mais interessante.Se o nosso objetivo imediato é analisar os elememtos tarsais dos Dasyuroidea, devemos também ter em mãos Dromiciops devido a seu posicionamento nesse conjunto, como grupo-irmão dos Didelphimorfia.
Entre os Dasyuroidea sem dúvida alguma o gênero mais conservador é Eudromicia. Esse gênero é plesiomórfico para quase todos os caracteres, mas com alguns avanços na pata (que é sindáctila com modificações nos ossos) e no crânio (principalmente nas zonas relacionadas diretamente às modificações mandibulares). Tendo em vista esses avanços não há muito com que se preocupar.
Apesar das modificações da pata ainda assim é o gênero menos modificado se o compararmos com os grupos externos.
Na figura vemos a ordem mais adequada para comparação: Sinodelphys, Caenolestes, Marmosa, Eudromicia e Dromiciops.
Em Marmosa o astrágalo possui três áreas (facetas) bem definidas em vista plantar : Calcâneo-astragalar (CaA), Sustentacular (Su) e astrágalo-navicular (AN) em verde na figura.
Em Sinodelphys e Caenolestes Su e AN são contínuas e separadas de CaA.
Em Dromiciops e nos Dasyuroidea SU e CaA estão unidas (contínuas).
Em Dasyuridae apenas Thylacinus é bem diferente e na base Murexia e Phascogale são difíceis de polarizar.
Em Peramelidae temos duas formas derivadas bem definidas, dois caminhos evolutivos diferentes.
Na ilustração acima utilizei desenhos de Szalay e Zhe-Xi Luo.
-Szalay, F. S. 1994. Evolutionary History of the Marsupials and an Analysis of Osteological Characters. New York: Cambridge University Press, 1995. 481p.
-Luo, Zhe-Xi; Ji, Qiang; Wible, John R.; Yuan, Chong-Xi (2003-12-12). "An early Cretaceous tribosphenic mammal and metatherian evolution". Science 302 (5652): 1934–1940.
What are the prerequisites needed to analyze the tarsal elements of the Dasyuroidea?............
In Dasyuridae only Thylacinus is quite different and the basal Murexia and Phascogale are difficult to polarize.
In Peramelidae have two well-defined derivative forms, two different evolutionary paths.
Na ilustração acima utilizei desenhos de Szalay e Zhe-Xi Luo.
-Szalay, F. S. 1994. Evolutionary History of the Marsupials and an Analysis of Osteological Characters. New York: Cambridge University Press, 1995. 481p.
-Luo, Zhe-Xi; Ji, Qiang; Wible, John R.; Yuan, Chong-Xi (2003-12-12). "An early Cretaceous tribosphenic mammal and metatherian evolution". Science 302 (5652): 1934–1940.
What are the prerequisites needed to analyze the tarsal elements of the Dasyuroidea?............
In Dasyuridae only Thylacinus is quite different and the basal Murexia and Phascogale are difficult to polarize.
In Peramelidae have two well-defined derivative forms, two different evolutionary paths.

Em baixo evolução de Pseudocheirus e Trichosurus, até Phascolarctos e Vombatus. Elementos tarsais de arboricolas até fossorial.
A polarização é marcante, Hypsiprimnodon retem mais o padrão ancestral que os demais macropodóides, e isso vai se juntar a característica mais importante ( presença do artelho 1 ). Prionotemnus é mais avançado e não encontra nenhum paralelo fora de seu grupo. Uma pequena faceta proximal calcâneo-astrágalo (adicional), também caracteriza esse grupo [em laranja].
O pequeno avanço de Phascolarctos em relação a Trichosurus é orientador sobre sua posição mais derivada a partir de uma forma semi-fossorial.
Vejam o paralelismo com os Peramelidae. Varias figuras aqui nesse álbum abordam esse tema.
O calcâneo de Marmosa tem uma característica "plesio" e outra "apo."
A forma geral do calcâneo é plesiomórfica, mas a condição da distância das facetas, APO.
-Em Marmosa o astrágalo possui três áreas (facetas) bem definidas em vista plantar : Calcâneo-astragalar (CaA), Sustentacular (Su) e astrágalo-navicular (AN) em verde na figura.
Entendimento é essencial. A coisa não é tão fácil assim.
Evolução dos Elementos Carpais em Marsupialia

Cefalização como forma de evolução arborícola. Apesar do ancestral dos marsupiais já apresentar transformações para a vida nas árvores (arborícola), existem graus de adaptação. Como uma cauda preênsil, que existe nos Didelphoidea, mas não nos Caenolestoidea.



O calcâneo de Marmosa tem uma característica "plesio" e outra "apo."
A forma geral do calcâneo é plesiomórfica, mas a condição da distância das facetas, APO.
-Em Marmosa o astrágalo possui três áreas (facetas) bem definidas em vista plantar : Calcâneo-astragalar (CaA), Sustentacular (Su) e astrágalo-navicular (AN) em verde na figura.
Entendimento é essencial. A coisa não é tão fácil assim.
Evolução dos Elementos Carpais em Marsupialia
 |
| Evolução dos Elementos Carpais - Marsupialia unciforme = hamate; magnum = capitate |
As principais modificações, feitas no trabalho de Szalay & Sargis 2001, foram: a posição relativa de Dromiciops e a resolução da dúvida em relação a posição de Notoryctes. Mas foram feitas outras observações.
O posicionamento proposital das figuras auxilia muito na visualização, ordenação das principais linhas e deslocamento das modificações...como fiz com Dromiciops para ressaltar.
Por uma questão de parcimônia temos que assumir que a modificação do magnum (capitate) ocorreu independentemente em Dromiciops, Dasyuride e Peramelidae, nesses dois últimos se explicaria muito bem como o resultado de uma locomoção terrestre. No caso de Dromiciops foi uma modificação maior que envolveu outros ossos resultando num estreitamento do pulso, talvez permitindo maior mobilidade.
Em Dasyuridae a modificação foi muito sutil, mas resultou num contato entre o unciforme (hamate) e o escafóide, talvez aumentando a rigidez da pata.
Szalay & Sargis publicaram essas ideias em 2001, eu já havia sugerido as relações filogéneticas de Notoryctes com os Peramelidae em 1991 em minha dissertação de mestrado, mostrando a evolução das patas nos marsupiais.
Este quadro não é simplesmente uma cópia....ele é claro, bem definido, sua disposição é lúcida e didática...e trás importantes mudanças e definições, além de ser atualizado e integrado."Uma das características mais significativas é o desenvolvimento do RADIO em Didelphimorfia".
Nos Didelphoidea e nos Dasyuroidea o radio é um osso mais forte e desenvolvido que nos ancestrais asiáticos, que nos Caenolestidae e mais desenvolvido que nos Microbiotheria, ou seja, temos aqui um caráter que só apareceu mais tarde certamente, pois até nos eutheria o radio é menos desenvolvido.
Como Microbiotheria compartilha 3 importantes características com os Didelphimorfia, ou seja, cauda preensil, artelho sem unha e calcanhar modificado, vários caracteres num conjunto integrado (muito forte). Fica mais evidente que os Didelphimorfia formam um grupo monofilético e a questão das facetas fundidas perde força diante do i3 encavalado (distribuição em mosaico).
Em associação com esse caráter temos o unciforme (hamate) que também se desenvolveu mudando o "design" da pata. Portanto, acompanhando as modificações da cauda e do pé na transição de Caenolestidae para os demais marsupiais atuais, houve uma modificação na mão que determinou dois rumos evolutivos. Microbiotheria e Didelphimorfia.
Diante desse quadro acima ficam fortalecidas as posições dos Caenolestidae, dos Microbiotheria e de Notoryctes, pois novas evidencias vêm confirmar essas relações. Isso é importante pois uma filogenia consistente nos permitirá compreender melhor sua história biogeográfica, e consequentemente a história do nosso planeta.
Didelphoidea Evolution
Os penises de Caluromys e Micoureus possuem a abertura externa da uretra localizada bem na ponta do órgão. Estes gêneros também possuem glândulas produtoras de mucos bem desenvolvidas. Eu considero essas características como sinapomorfias de Caluromys e Micoureus, uma adaptação à vida arbórea. A cópula desses animais ocorre num ambiente onde as opções de posicionamento são bastantes limitadas. Por isso um penis mais alongado com abertura na ponta e muita muita mucosidade ajuda na transferência dos gametas. Em Marmosa temos uma situação um pouco mais conservadora com relação à abertura da uretra. Comparem com Gracilinanus e Didelphis. Caluromys e Micoureus também possuem um pelo mais lanoso e a cauda bem longa comparativamente com pintas escuras na base. Possuem um crânio com características derivadas como processos pós-orbitais desenvolvidos e focinho encurtado. Caluromys é um extremo evolutivo entre os Dedelphoidea. Assim como Chironectes.
As relações estreitas existentes entre os gêneros Caluromys e Micoureus são tão óbvias para mim, que chego a dar risadas, quando leio certos trabalhos colocando esses gêneros em posições tão estranhas e variadas ! Está muito claro para mim que Micoureus é um passo à frente em relação à Marmosa, e é o grupo irmão dos chamados "Caluromyinae". Eu considero Marmosa + Tlacuatzin + Micoureus + Caluromys + Glironia + Hyladelphys...como Marmosinae uma subfamília de Marmosidae. "Caluromysiops" por uma questão de monofilia é um Caluromys !

Comparação entre Monodelphinae e Marmosidae.
Cefalização como forma de evolução arborícola. Apesar do ancestral dos marsupiais já apresentar transformações para a vida nas árvores (arborícola), existem graus de adaptação. Como uma cauda preênsil, que existe nos Didelphoidea, mas não nos Caenolestoidea.
Didelphoidea = Marmosidae + Didelphidae
Marmosidae = Marmosinae + Thylamyinae Didelphidae = Monodelphinae + Didelphinae

 |
| Arboreal life adaptations in skulls of Mammals - Round brain cases, Shorted muzzles, Orbital orientation: frontal, Higher mandibles with angular process expanded and Reductions in teeth size or number |


The evolution of the mandible. Adaptations to an arboreal life. Micoureus, Caluromys and Glironia.
Desdobramento progressivo do processo angular da mandíbula.
Desdobramento progressivo do processo angular da mandíbula.
Muito bom esse texto. Mostra uma perspectiva filogenética a partir dos grupos atuais. Parabéns.
ResponderExcluirObrigado pela sua presença Édison !
ResponderExcluirOs grupos atuais devem necessariamente centralizar a pesquisa sobre os Mammalia ou qualquer outro grupo, pois podem ser estudados na íntegra.
Os fósseis são muito importantes, mas são interpretados sempre por comparação com os viventes.
Existe uma certa ordem nos estudos gerais. O mesmo vale para a genética. Não adianta comparar genes sem conhecer suas correspondências fenotípicas.
Mas esse tempo ainda não chegou. A mentalidade tipológica ainda domina a cena. Grande parte dos biólogos ainda se debate entre tipologia e evolucionismo.
Vivemos um tempo caótico depois da "onda" molecular.
Os tipologistas pensam que estão falando de evolução, mas estão completamente enganados.