Aqui, o conceito de EUTHERIA se apresenta de uma forma diferente daquela que normalmente se define por esse termo. Como já apareceu anteriormente, os PRETHERIA (Xenarthra) e os LATHERIA (Pholidota), que normalmente são considerados como parte dos Eutheria, aqui estão separados destes. Adicionalmente a isso, eu divido os EUTHERIA em ARBOREA e TERRESTRIA. Diferentemente do que tem sido estabelecido ao longo do tempo, ou seja, na história taxonômica dos Mammalia, aqui considero os METATHERIA (Marsupialia) como o grupo irmão imediato dos EUTHERIA, que juntos formam os THERIA, tendo como grupo irmão imediato os LATHERIA (Pholidota), os pangolins, e como grupo basal final os PRETHERIA (Xenarthra). Finalmente, todos eles juntos formam os PLACENTALIA. ver Fig.
1 - P R O T O T H E R I A (MONOTREMATA)
2 - P L A C E N T A L I A (VIVIPARA)
- PRETHERIA (XENARTHRA)
- LATHERIA (PHOLIDOTA)
- THERIA
I - METATHERIA (MARSUPIALIA) [Caenolestoidea, Microbiotherioidea, Didelphoidea, Dasyuroidea]
II - EUTHERIA
1) ARBOREA
- Archonta (living) [Scandentia, Primates, Dermoptera, Chiroptera]
- Ferae (living) [Carnivora]
- Ungulata (living) [Artiodactyla, Perissodactyla]
2) TERRESTRIA
- Zalambdodonta (living)
- Ferae (living) [Carnivora]
- Ungulata (living) [Artiodactyla, Perissodactyla]
2) TERRESTRIA
- Zalambdodonta (living)
[Solenodontoidea, Chrysochloroidea, Tenrecoidea]
- Hypoconodonta (living)
- Hypoconodonta (living)
[Erinaceoidea] (Erinaceidae - Soricidae - Talpidae)
[Caecoidea] (Tubulidentata - Macroscelidea - Peanungulata - Glires)
[Caecoidea] (Tubulidentata - Macroscelidea - Peanungulata - Glires)
 |
| Cladograma da Evolução das Ordens dos EUTHERIA, mais o grupo irmão METATHERIA (Marsupíalia), ou seja os THERIA. |


A EVOLUÇÃO do CRÂNIO NOS MAMMALIA
 |
Nos ARBOREA as transformações cranianas, resultantes das adaptações à vida arborícola, produziram o desenvolvimento do encéfalo, como resultado da curvatura do crânio. Houve também um incremento da visão, em detrimento da olfação, provocado pela redução do focinho. Mas a cefalização atingiu o seu grau máximo entre os ARCHONTA: pois os Ferungulata basais podem apresentar uma cefalização próxima da ancestral, um pouco mais avançados que os MARSUPIALIA. Alguns TERRESTRIA também apresentam crânios de formas arredondadas, porém nitidamente pedomórficos, como Megasorex, Galemys, Microgale e Amblysomus hottentotus, todos com características fetais como olhos muito reduzidos e arcos zigomáticos delicados ou ausentes. Lista de Táxons da Figura acima: ARBOREA: 1.(Hemigalus derbyanus) Viverridae, Carnivora, Ferae. 2.(Acerodon jubatus) Pteropodidae, Chiroptera, Archonta. 3.(Galeopterus variegatus) Dermoptera. Archonta. 4.(Tupaia glis) Tupaidae, Scandentia, Archonta. 5.(Microcebus rufus) Lemuridae, Primates. Archonta. TERRESTRIA: 1.(Echinosorex gymnura) Erinaceidae, Hypoconodonta. 2.(Megasorex gigas) Soricidae, Hypoconodonta. 3.(Galemys pyrenaicus) Talpidae, Hypoconodonta. 4.(Paramys delicatus) Rodentia, Hypoconodonta. 5.(Petrodromus tetradactylus) Macroscelidea, Hypoconodonta. 6.(Orycteropus afer) Tubulidentata, Hypoconodonta. 7.(Solenodon paradoxus) Solenodontidae, Zalambdodonta. 8.(Potamogale velox) Potamogalidae, Zalambdodonta. 9.(Amblysomus hottentotus) Chrysochloridae, Zalambdodonta. 10.(Tenrec ecaudatus) Tenrecidae, Zalambdodonta 11.(Microgale cowani) Tenrecidae, Zalambdodonta. Os TERRESTRIA retiveram um crânio achatado e uma olfação muito desenvolvida, ou seja, a condição reptiliana. Os olhos pequenos se reduziram ainda mais em certas linhas evolutivas como nos Chrysochloridae, que não possuem olhos externos. Essa redução também é bem significativa entre os pedomórficos Talpidae e Soricidae. A evolução dos Terrestria é sem dúvida marcada pelo desenvolvimento da olfação, pela vida fossorial e o desenvolvimento de uma probóscis (uma tromba), que representa a principal característica sinapomórfica desse grupo. Geralmente suas vibrissas são numerosas e desenvolvidas, apresentando correspondências significativas no encéfalo. Obviamente esses caracteres citados são basais, se apresentando muito modificados nos ramos mais derivados. como nos elefantes, peixes-bois e em outros Caecoidea (o grupo mais avançado). |
 |
| A EVOLUÇÃO do ENCÉFALO dos MAMMALIA Essa sem dúvida é a melhor figura para compararmos o grau de cefalização nos EUTHERIA. Existem alguns símbolos facilmente compreensíveis indicando a curvatura do encéfalo, região do mesencéfalo e o ângulo do avanço do neocórtex sobre o cerebelo. O desenvolvimento dos lóbulos temporais é marcante nos ARBOREA. Além do foco especial em cima da cefalização podemos ver também os resultados da pedomorfose, que marcaram toda a evolução dos Talpidae-Soricidae. Nesse grupo o encéfalo adquiriu uma forma globosa nos Soricidae (veja Blarina) e se simplificou. O tamanho e a complexidade do encéfalo estão relacionados principalmente com o tamanho do animal. Animais grandes, como elefantes e cetáceos, apresentam encéfalos grandes e complexos, isso se deve a quantidade de receptores de sensibilidade e suas relações motoras correspondentes. Animais muito pequenos, como musaranhos e roedores, possuem encefálos mais lisos e reduzidos. |
 |
| A EVOLUÇÃO do SCROTUM. Nos Mammalia observamos escrotos de 2 tipos: pré-peniano e pós-peniano. |
O scrotum (ou escroto) é uma característica que aparece apenas em alguns mamíferos. Os Marsupialia se caracterizam por possuírem um escroto pré-peniano pedunculado, situado cranialmente à cloaca, uma estrutura exclusiva deles (sinapomórfica). Existe uma condição algo semelhante nos Lagomorpha, que também possuem uma cloaca, com os testículos situados cranialmente ao penis (mas não possuem um escroto verdadeiro). Além de uma cloaca, vários gêneros de marsupiais apresentam a glande bifurcada, como nos ornitorrincos, uma herança reptiliana.
Grande parte dos ARBOREA apresenta um escroto verdadeiro, mas esse caráter foi perdido nos cetáceos e em outros membros desse grupo. Sem dúvida essa característica é sinapomófica para todos os ARBOREA. Geralmente o escroto é pós-peniano, mas pode ser lateral em alguns membros basais de Archonta. Nenhum ARBOREA possui uma cloaca, e isso certamente está relacionado a aquisição de um escroto pós-peniano, pois são estruturas incompatíveis. Os marsupiais conservaram a cloaca, mas desenvolveram um escroto pré-peniano pedunculado.
O ancestral de todos os TERRESTRIA possuía uma cloaca pré-caudal, que ainda persiste em muitos deles. Todos os Zalambdodonta (exceto Solenodon) possuem uma cloaca e os testículos são abdominais. Nenhum dos Terrestria apresenta um escroto verdadeiro. Outros grupos de Terrestria atestam a existência de uma cloaca no ancestral comum. Todos os Lagomorpha e dois membros basais de Rodentia possuem cloaca: Aplodontia e Castor. Também foram encontradas cloacas em vários Soricidae e Erinaceidae. Isso significa que os dois ramos principais dos Hypoconodonta ainda carregavam esse caráter.
"Entre as características primitivas que muitos insetívoros exibem estão a estrutura das orelhas, o cérebro pequeno, os dentes primitivos, os testículos que geralmente estão dentro do abdômen, e não no escroto, e a união dos tratos urinário e reprodutivo e do intestino em um canal comum chamado cloaca. Os musaranhos, mais primitivos que os roedores, têm um cérebro menor e, como os répteis, muitas espécies, incluindo o musaranho de cauda curta do norte (Blarina brevicauda) e o musaranho menor (Cryptotis parvus), têm os tratos genital e urinário fundidos em uma única abertura. chamada de cloaca". Site Appalachian Mammals, de Bob Pickett.
Outros Eutheria também possuem cloacas: os Tenrecidae e os Chrysochloridae. Os gêneros Aplodontia e Castor. Muitos Soricidae, Potamogale e Ochotona. Todos os Lagomorpha possuem uma cloaca. Como o canal inguinal de lagomorfos e roedores permanece aberto durante toda a vida, não existe um escroto verdadeiro, mas sim dois sacos hemiescrotais separados. Nos lagomorfos, eles são posicionados cranialmente ao pênis; nas espécies de roedores, são caudais ao pênis, assim como em outras espécies de mamíferos placentários (exceto coelhos). O termo “canal” não é apropriado para roedores, porque é muito largo. Os sacos hemiescrotais apresentam peculiaridades anatômicas entre diferentes grupos de roedores. Roedores semelhantes a ratos (o rato comum, o camundongo, o hamster e os gerbils) têm sacos hemiescrotais bem desenvolvidos. Não há um escroto verdadeiro (Brower, 2006; Donnelly and Brown, 2004; Hoefer and Crossley, 2002).
Richard Owen in 1863:57 ao reconhecer a identidade primata do Aye-Aye (Daubentonia madagascariensis), nos ilumina sobre a origem dos roedores, num texto muito útil para os zoólogos gradistas atuais:
“Os órgãos de geração são importantes indícios de afinidade natural na classe dos mamíferos, mais especialmente os órgãos masculinos, dos quais o sexo do Aye-aye por mim dissecado permite esta aplicação. Em todos os Roedores a abertura peritoneal do saco seroso dos testículos é tão largo, e o cremaster tão grande e tão disposto, que a glândula pode ser retirada para o abdômen, e emerge em um escroto séssil temporário apenas sob o aumento sazonal para procrição, o que é considerável. Assim, os Rodentia são ‘testiconda temporários’. O Aye-aye não tem esta organização: os testículos são permanentemente ‘escrotais’ após sua passagem para fora do abdômen. Os esquilos, como outros roedores, têm “vesículas seminais” distintas com espessas camadas glandulares: nos Leporidae a vesícula é única, mas grande. A ausência de bolsa ou bolsas vesiculares distintas no Aye-aye o remove dos Rodentia, enquanto o tamanho e a forma da próstata* e das glândulas de Cowper o aproximam dos Lemuridae.
Um testemunho mais decisivo é dado pelo pênis. Os esquilos e todos os Rodentia são 'retromingentes', o pênis sendo curvado repentinamente sobre si mesmo, com a 'glande' situada em um prepúcio que se abre perto do ânus. No Aye-aye, como nos Lemuridae e em outros Quadrumana, a bainha prepucial do pênis se projeta livremente para a frente na medida indicada pelo caráter lineano 'pendular', aplicado aos 'Primatas' de modo geral no 'Systema Nature' ".
OWEN, RICHARD. (1863). Monograph on the Aye-aye Chiromys madagascariensis, Cuvier. London, Taylor and Francis, 1863. 72 pp.
________________________________________________________________________
ARBOREA (Gomes, 2015) : Superorder ARCHONTA (Gregory, 1910), Volitantia (Illiger, 1811)
VOLITANTIA: Dermoptera + Chiroptera
 |
| Observe que Dermoptera tem um crânio mais derivado, apesar de todos os seus caracteres plesiomórficos. |
Este fóssil do começo do Eoceno : Onychonycteris finneyi, é o mais antigo fóssil de Chiroptera conhecido. Onychonycteris significa "morcego de garras", pois ele é o único morcego conhecido que tem 5 garras ou seja em todos os dedos. Os Pteropodidae tem garras nos dedos 1 e 2, e os Microchiroptera só no dedo 1. Temos aí uma série de transformação.
Os Pteropodidae ainda retém os membros posteriores muito grandes semelhantes aos de Dermoptera. Pedomorfoses sucessivas ao longo da evolução dos Chiroptera, moldaram animais de pequeno porte com crânios cada vez mais modificados próximos de formas fetais.
Os Pteropodidae ainda retém os membros posteriores muito grandes semelhantes aos de Dermoptera. Pedomorfoses sucessivas ao longo da evolução dos Chiroptera, moldaram animais de pequeno porte com crânios cada vez mais modificados próximos de formas fetais.
A fórmula dentária de Onychonycteris é exatamente a esperada para o ancestral dos Chiroptera:
2/3, 1/1, 3/3, 3/3 = 38
Dentre os grupos incluídos nos Archonta, os Dermoptera e os Chiroptera seguramente formam um grupo monofilético já reconhecido e denominado como Volitantia. Porém as relações existentes entre os Primates e os Volitantia, ou entre estes e os Scandentia, indicam uma "tricotomia legítima". Pois existem caracteres derivados compartilhados, que justificam uma derivação comum entre Primates e Volitantia, principalmente referentes à visão e regiões cerebrais relacionadas. Também a glande do pênis formada por uma extensão do "corpus spongiosum" é uma sinapomorfia que reune Primatas, Dermoptera e Chiroptera [esse caráter é universalmente exclusivo deles]. Por outro lado, também existem caracteres derivados que são exclusivos dos Volitantia e dos Scandentia, como costelas achatadas, uma característica também encontrada em Ptilocercus. Essa relação com Ptilocercus não para por aí, a escápula é flutuante e alta, as vértebras cervicais largas, "a mão também é muito semelhante em dois aspectos", o carpo é rotativo e as proporções metacarpo/falanges muito semelhantes.
 |
| ARBOREA (Gomes, 2015): ARCHONTA (Gregory, 1910) |
Considero essa situação como uma distribuição em mosaico que se explica melhor por uma verdadeira tricotomia. Que não deve ser "resolvida"...mas apenas compreendida. Nesse conjunto certamente os Scandentia representam um grupo mais conservador, mas isso não justifica a união dos demais. Pois os Primates formam um agrupamento bem diversificado com muitas derivações próprias, assim como os Chiroptera. No entanto, por possuírem um (anel orbital fechado por osso), os Scandentia já foram considerados primatas.
Vale a pena lembrar que os Microchiroptera passaram por uma evolução pedomórfica, perdendo muitas das características que são comuns entre os Pteropodidae. Isso levou muitos pesquisadores a questionar a união desses dois grupos, mas isso é um grave erro metodológico, fruto de análises gradistas com pretensões cladísticas, que infestam a zoologia.
Nas últimas décadas a cladística foi totalmente deturpada pela FENÉTICA, nos levando a esse caos que está aí. Indução embrionária e alometria......pedomorfose como uma forma rápida de evolução....sua importância adaptativa.....e particularmente a regressão de caracteres....põem a baixo uma série de metodologias comparativas como "a FENÉTICA e a MOLECULAR".
_________________________________________________________________________________________
Note that the skull is more derived in Dermoptera, despite all its plesiomorphic characters.
This fossil of the early Eocene: Onychonycteris finneyi, is the oldest known fossil of Chiroptera. Onychonycteris means "clawed bat" because it is the only known bat that has 5 claws i.e. in all fingers. The Pteropodidae has clawed fingers 1 and 2, and Microchiroptera has only on the finger 1. Here we have a series of transformations.

The Pteropodidae still retains the very large hind limbs similar to Dermoptera. Successive pedomorfosis along the evolution of Chiroptera, molded small animals with skulls increasingly modified near fetal forms.
The dental formula in Onychonycteris is exactly the expected to ancestral Chiroptera:
2/3, 1/1, 3/3, 3/3 = 38
Embryonic induction and allometry...paedomorphosis as a quick way of evolution...its adaptive significance...and particularly regression of characters...puts down a series of comparative methodologies such as "the Phenetics and the MOLECULAR "
__________________________________________________________________________
Em 1811, Illiger criou os táxones Volitantia e Dermoptera. Volitantia foi tratada como Ordo XI. e Dermoptera como Familia 31. (I). Volitantia reunia os Chiroptera e os Dermoptera, estes últimos, eram representados pela única espécie conhecida na época: Galeopithecus volans, baseada no Lemur volans de Linnaeus, que a colocou dentro da Ordem PRIMATES, a qual abrigava também os morcegos: Genus Vespertilio. O táxon Chiroptera foi criado por Johann F. Blumenbach (1779:74), e incluía apenas cinco espécies colocadas no gênero Vespertilio. Em 1875, George E. Dobson divide a Ordem dos Chiroptera em duas subordens : Megachiroptera e Microchiroptera.
V O L I T A N T I A (Illiger, 1811)
Linnaeus, (1758: 30), Systema Naturae.
Cato- Simius volans camelli. Pet. gaz. 14. t. 9. f. 8.
Act. angl. 277. n. 1065.
Vespertilio admirabilis. Bont. jav. 68. t. 69.
Felis volans ternatea. Seb. mus. 1. p. 93. t. 58. f. 2. 3.
Habitat in Asia.
Membrana more Sciuri volantis s. Muris volantis a ca-
pite ad manus, a manibus per latera ad plantas, a
plantis ad apicem cauda expansa. Ungues acuti.
Mammae duae pectorales. Edit fructus arborum. Ex
mammis videtur Lemuribus s. Simiis proximum A-
nimalculum, sed nobis non visum; ulterius examinent
autoptae.
[Lemur] voador. 3. Lemur caudado, membrana voadora circundante. †.
Gato-Macaco voador G. J. Kamel. J. Petiver. Gazophylacium. 14. tabula. 9. fig. 8. (S.B.6.I.) & Philosophical transactions R. S. L. n. 277. p. 1065.
Gato voador tripartido. Seba. museu. I. p. 93. tabula. 58. figs. 2. 3.
Ela vive na Ásia
Membranas à maneira de um esquilo voador ou
de um rato voador, estendendo-se da cabeça às mãos,
das mãos pelos lados até as plantas dos pés, das plantas
até a ponta da cauda. Garras agudas.
Duas mamas peitorais. Ela come os frutos das árvores.
De acordo com as mamas, parece ser um pequeno animal próximo aos Lêmures ou aos Macacos, mas não o
examinamos; serão necessários mais exames nos espécimes .
Linnaeus descreveu o Lemur Voador a partir de três referências da literatura, citadas como sinônimos. A mais antiga delas é a de Jacobus Bontius, que foi publicada em 1658. Ele foi o primeiro a descrever esse animal, ao qual ele se referiu como Vespertilio admirabilis, ou seja, o Morcego admirável. Leia na íntegra, logo abaixo, a sua detalhada descrição, a qual foi publicada depois de sua morte por Guilherme Piso.
CAPITULO XVI
Apêndice. Do Morcego admirável.
Fica especialmente claro a partir desses morcegos, que não só a África, mas também a Ásia e a América estão cheias de monstros, como os muitos mencionados acima; Pois em Gusaratta, sob o Império dos Grandes Mongóis, eles voam em bandos como gansos selvagens, e ao entardecer são vistos suspensos no ar ou em árvores, e deixam todos os visitantes surpresos com seu tamanho incomum, volume e conformação estranha. Assim, os belgas, hesitando com a novidade do assunto, se deveriam atribuí-lo ao gênero das aves, deram o nome um tanto incongruente de macacos alados. Que eles usam barbatanas membranosas em vez de asas, que são igualmente douradas, e que são providos de garras e quatro patas, e que a filhotes, e não a ovos, eles dão à luz e amamentam, devem ser atribuídos às famílias dos outros Vespertilianos: É verdade que têm o tamanho dos gatos, com a barriga e o peito grossos e carnudos, e também que são cobertos por uma membrana contínua como um véu expandido do pescoço e da cabeça até a ponta da garra, pois parecem diferir do resto dos morcegos. Acrescente-se a isso que o véu por baixo é realmente membranoso, de modo que parece coberto por pelos, penugem, veias e fibras, mas por fora ele é revestido de uma maneira estranha, com uma pelagem permanente mais macia, semelhante à dos coelhos, e de pelos pretos e grisalhos. Também aquelas pregas, pelas quais as asas, à maneira dos outros morcegos, são contraídas e relaxadas, o deixará desapontado. De modo que, se você comparar com o aspecto do morcego brasileiro, ou de qualquer outro semelhante, você o achará muito diferente deles. Mas, para que o leitor curioso não sinta a falta, exibo duas figuras, uma voando, e outra apoiada nas costas e estendida. O animal inteiro tem cerca de três pés de comprimento e o mesmo de largura. A cauda é fina, a extensão é longa e ela adere à membrana de todo o corpo, como se estivesse imersa. As pernas também são cobertas por uma penugem amarelada mais suave. Das últimas juntas dos pés saem cinco garras finas, muito afiadas e curvas, com as quais o mordedor segura tudo o que apanha, principalmente os frutos das árvores, que destrói, feitos de uma película muito fina.
Eu poderia acrescentar mais à disposição e geração desses vespertílios, que de fato foram fornecidos a mim por nossos antigos; mas como eles estavam dispersos e pareciam ser relatórios nublados, preferi me deter em sua descrição, até que a verdade fosse mais apurada, para que, ao preencher as páginas, erros (que costumam acontecer em anexos exóticos) fossem propagados.
Além disso, segundo Bontius, nossa testemunha, morcegos também são comuns em Java, vivendo na luz, do tamanho de pombas, dos quais os javaneses se alimentam, afirmando que são muito saborosos. Às vezes, à noite, eles entram nas camas, seja por causa do calor do dia, ou se espelhos ou janelas foram deixados abertos, e mordem os pés descalços dos que dormem, tirando uma grande quantidade de sangue, com mais medo do que perigo, para aqueles que compartilham essas coisas.
* * *
Em 1702, James Petiver, na Gazophylacium naturae et artis, baseado no relatório do jesuita Georg Joseph Kamel, registrou o seguinte : "8. Cato Simius volans Cameli. Veja sua descrição na Philosophical Transactions nº 277. pág. 1065. Este estranho animal é encontrado nas ilhas filipinas, e sua morada habitual é nas árvores".
Philosophical Transactions. (Number. 277.) para os meses de janeiro e fevereiro, 1702.
Aqui seguem alguns animais, &c. Observados nas Ilhas Filipinas, por aquele reverendo e erudito padre George Joseph Camel, de quem recentemente os recebi.
2. Cato-Simius volans Cameli. gazophyl. nost, Naturae Tab. 9. fig. 8.
Colago e Cagvang em Bysaiani [Ilhas Visayas], Gigua em Pampangi e Tagali. [idiomas filipenses].
Esta estranha e maravilhosa criatura tem o tamanho de um gato, seu corpo é como o de um macaco, mas mais esguio; da cabeça à cauda tem 3 palmos, entre os braços estendidos 2 e entre as coxas 1 e meio. São três palmos da frente para os dedos traseiros e outros tantos entre as 2 patas dianteiras, mas entre os traseiros são apenas 2 palmos. Sua barriga tem a largura de uma mão e a pele dela um palmo. Eles relatam que na província de Pampanga, eles são tão grandes quanto um guarda-chuva chinês, ou 6 palmos de largura.
Seu pelo é castanho, na parte de trás de um tom cinza-rato, misturado com um branco muito curioso, despojado, como aquelas peles que os alemães chamam de feck (que talvez seja nosso fitchow ou furão), estas são mais longas no corpo e mais curtas em sua pele membranosa. Essa pele é estendida ao redor de seu corpo da cabeça à cauda como um morcego, com a qual, sendo expandida, eles lentamente vão do topo de uma árvore para o meio da próxima, e então avançam por ela dando saltos.
Ele tem cara de macaco, orelhas nuas redondas de morcego, cada pé tem 5 garras achatadas e afiadas como as de um leão: na barriga das fêmeas há 2 saquinhos, aos quais os filhotes se agarram e chupam: não sabemos qual é sua comida, mas provavelmente é fruta, porque a sua morada habitual é nas árvores. Veja na figura, que o padre Camel me enviou, na Tabula 9 do meu Gazophylacium naturae.
* * *
Em 1734, Albertus Seba descreveu na página 93 do volume I do seu Tesauro, um animal que ele denominou como Felis volant, Ternatana; fœmina (Gato voador tripartido, fêmea). Logo em seguida, ele também descreveu um macho. Aqui apresentarei apenas a descrição da fêmea.
N°. 2. Gato Voador Tipartido, fêmea.
Este animal difere totalmente do anterior, pois suas asas, tanto externa quanto internamente, são cobertas por pelos finos e macios, como as toupeiras, de cor cinza-avermelhada. Sua cabeça parece mais com a de um gato selvagem do que de um doméstico, com focinho de cachorro. Suas orelhas são de comprimento medíocre: seus olhos grandes e brilhantes; suas asas peludas se estendem de maneira diferente da espécie anterior. Suas quatro patas terminam em unhas curtas e curvas, e junto com a cauda, são ampliadas pelo tecido membranoso das asas. Dessa forma, todas as partes a ajudam a voar, inclusive a cabeça, que se une a elas por meio das asas, que se projetam até o pescoço e cruzam sob o queixo por meio de uma manta. Sua cabeça, grande e redonda indica que é uma fêmea. Não deixe de notar que o tecido membranoso das asas parece ter um contorno serrilhado, e que sua superfície interna é menos peluda, e de um amarelo menos escuro, que a externa. Voa muito rápido, mas baixo, de modo semelhante aos nossos morcegos, andando em círculos em busca de alimento.
_____________________________________________________________________________
XI. Ordo.
Volitantia (Flatterfüsser.)
- Dentes superiores da frente de 2-4, inferiores de 2-6, alguns decíduos. Dentes cortadores, nos Dermópteros ambíguos, nos Chirópteros distintos. Molares recobertos com cúspides.
- Duas mamas abertas, peitorais. Genitália distinta do ânus.
- Pés virados para fora, Dermóptero ou Chiróptero, patágio no tronco e circundando o ânus. Unhas falciformes.
- Nome. Esta ordem inclui apenas mamíferos realmente voadores, de onde vem a sua denominação.
I. DERMOPTERA [ Illiger, 1811: 116 ]
31. (I) Familia.
Dermoptera (Pelzflattrer.)
- Dentes da frente de ambos os lados 4, inferiores pectinados. Dentes cortadores ambíguos. Molares cobertos com cúspides.
- Duas mamas abertas, peitorais.
- Pés Dermópteros, patágio piloso lombar e anal.
dedos da mão juntos, formando um manículo, semelhante a um podário, conectado a aba.
GENUS 81. GALEOPITHECUS Pallas, La Cep. Geoffroy, Dinner. Shaw. Tiedem. Leraur Storr (Pelzflattrer Germ.
Galeopitheque Gall. Colugo Angl.)
- Dentes superiores da frente 4, separados, semelhantes aos molares; inferiores 4 procumbentes pectinados. Caninos ambíguos, semelhantes aos molares proximamente relacionados a eles. 5 molares cobertos acima de cada lado, o primeiro dos quais é serrilhado como o canino superior, os 4 molares posteriores são cuspidados; em baixo 6 molares cuspidados.
- Focinho pontudo. Orelhas pequenas e arredondadas.
- Corpo circundado por um patágio piloso lombar conectando as mãos com as pernas, e um anal, conectando as pernas posteriomente.
- Cauda congenita com o curto patágio anal. 2 mamas peitorais abertas.
- Patas pentadáctilas com membranas, dedos da mão mais curtos que a ulna, juntos, não percorrendo o patágio como um raio, mas formando um manículo semelhante a um podário por fora do patágio, conectado com a aba. Garras falciformes muito comprimidas, laminares e agudas.
- Species: Lemur volans Lin.
Em 1869, o naturalista britânico Alfred Russel Wallace, em "The Malay Archipelago", escreveu: "Outro animal curioso, que encontrei em Cingapura e em Bornéu [...] é o Galeopithecus, ou lêmure voador. Esta criatura tem uma membrana larga que se estende ao redor de seu corpo até as extremidades dos dedos dos pés e até a ponta da cauda bastante longa. Isso permite que ele passe obliquamente pelo ar de uma árvore para outra…". Pelo texto do experimentado Wallace, fica claro, que na metade do século XIX, esses animais ainda eram pouco conhecidos pelos naturalistas europeus.
II. CHIROPTERA [ Blumenbach, 1779: 74 ]
Blumenbach, 1779. Handbuch der Naturgeschichte, p. 74.
V. CHIROPTERA.
Os dedos das patas anteriores são, excluindo o polegar, mais longos do que todo o corpo desses animais; e entre eles estende-se uma pele floral, que serve de asas (p.43). Portanto, eles podem andar tão pouco confortavelmente na terra, quanto os macacos.
II. V E S P E R T I L I O. morcego. Pollex palmarum et digiti plantarum breves, reliqui longissimi.
1. Spectrum. O vampiro. V. ecaudatus, naso infundibili formi lanceolato. *
Seba, thesaur. I. tabula LVIII. fig. 1.
Com as asas abertas, o corpo desta besta que habita a América Central é do tamanho do esquilo. Tem cor marrom-acinzentada, alimenta-se de animais e frutas, porém torna-se terrível porque suga o sangue de pessoas adormecidas, enquanto as atinge com suas grandes asas.
2. †. Perspicillatus. V. caudatus. naso folio simplici lanceolato. *
Exceto pela aba do nariz, eles são como o morcego comum. Na América do Sul, vivem nas casas e como sabemos, também na Alemanha. Vimos vários capturados em Gotha, em Jena, e em outras cidades.
3. †. Auritus. V. caudatus, auriculis maximis. *
Geralmente, mas de forma injusta, é atribuído a eles ter orelhas duplas. Elas são tão simples quanto os da espécie seguinte, no entanto, todas as partes (especialmente a concha com suas duas cristas e a folha frontal) são enormemente grandes, e por isso o animal tem um aspecto extremamente estranho. É comum na Europa e seu modo de vida é exatamente igual ao da espécie seguinte.
4. † Murinus. V. caudatus, auriculis capite minoribus. *
Esses animais se abrigam em paredes velhas e gostam de se alojar em despensas com toucinhos, porque eles se engancham com as garras dos polegares e os comem. À noite, e especialmente nas noites claras de verão, eles saem esvoaçantes, pegando mariposas, mas as próprias corujas podem pegá-los facilmente. Durante a hibernação, eles se penduram em grupos nas cavernas pelas patas traseiras.
5. Molossus. V. caudatus, auriculis crassis, brevibus, in front approximatis. *
No Brasil, habitam as casas, tem uma boca larga achatada, e uma dentição estranha. No espécime que temos diante de nós (Tabula. I. Fig. 4.), no maxilar superior existem dois dentes pontiagudos, no maxilar inferior existem dois dentes frontais extremamente pequenos e rombudos. Os dois caninos superiores são grandes e bem separados; os inferiores são um pouco menores, ficam mais próximos e estão na borda interna, em direção aos dentes da borda, um estranho e forte, que provavelmente alguns homens famosos tomaram por um par de dentes da borda. Em cada lado de cada maxilar existem quatro molares, dos quais os superiores são mais planos, os inferiores irregulares.
 |
| Molossus Tabula I. Fig. 4. (Figura de Blumenbach, modificada, restringida apenas para Molossus) |
* * *
A família Pteropodidae foi descrita pela primeira vez em 1821: 299 pelo zoólogo britânico John Edward Gray. Ele nomeou a família "Pteropidae" (baseado no gênero Pteropus) e a colocou dentro da agora extinta ordem Fructivorae. Fructivorae continha uma outra família, a agora extinta Cephalotidae, contendo um gênero, Cephalotes (agora reconhecido como sinônimo de Dobsonia). A ortografia de Gray foi possivelmente baseada em um mal-entendido do sufixo Pteropus. Pteropus vem do grego antigo pterón que significa "asa" e poús que significa "pé". A palavra grega pous de Pteropus vem da raiz pod-; portanto, latinizando Pteropus resulta corretamente no prefixo Pteropod-. O biólogo francês Charles Lucien Bonaparte foi o primeiro a usar a grafia corrigida Pteropodidae em 1838: 23. Wikipedia
JOHN EDWARD GRAY (1821)
On the Natural Arrangement of Vertebrose Animals
The London Medical Repository, Monthly Journal, and Review. Vol. XV. 1821.
Body horizontal, hairy : thumb separate on the upper
extremities only, clawed : fingers very long, interstices filled
up with a double nearly naked membrane, adhering to the
body and lower extremities, forming true wings: breast bone,
keeled : mamma two, pectoral, naked : teeth, three sorts,
cutting, canine, and grinders : penis free : coecum none.
Ord. 1. FRUCTIVORAE.

separated by a furrow : index finger short, of three phalanges:
eyes large, ears small: legs not united together.
from the sides of the body and legs: index finger with a
small claw : nose simple: tongue hairy : teeth, four cutting
in each jaw.
Pterope, Pteropus. Geoff. P. edulis. Geoff.
Roussette, Rousettus. Gray. P. AEgyptiacus. Geoff.
Fam. 2. CEPHALOTIDAE - Membrane of the wings
attached longitudinally to the middle of the back of the body,
and legs: index finger clawless: teeth, two cutting in each
jaw.
Cephalote, Cephalotes. Geoff. C. Peronii. Geoff.
Ord. 2. INSECTIVORAE.
Teeth, grinders, all acutely tubercular: index finger not
clawed: eyes small: ears large: legs united by a membrane.
Fam. 1. NOCTILIONIDAE - Middle finger with two bony
phalanges: the index and other fingers with only two bony
phalanges.
Race I. Nose simple: tail none, or adherent to the inter-
femoral membrane.
Mollosses. Geoff. Vespertilio mollossus. Gmel.
Nyctimones. Geoff. Vesp. acetabulosus. Herm.

Race 2. Nose warty: tail short, free.
Noctilio. Lin. Vespertilio leporinus. Gmel.
Race 3. Nose appendaged.
Vampyre, Phyllostoma, vespertilio spectrum. Lin.
Fam. 2. VESPERTILIONIDAE. - Index finger with only one
bony phalange ; the middle and other fingers, with two each.
Race 1. Nose appendiced.
Megadermes. Geoff. Vestertilio spasma Lin.
300 Original communications.
Leafnose, Rhynolophus. Geof. Vespertilio ferrum equi-
num. Lin.
Nyctere, Nycterus. Geof. Vespertilio hispidus. Lin.
Rhynopoma. Geoff. R. microphylla Geoff.
Thaphosores. Geoff.
Race 2. Nose simple.
Bat, Vespertilio. Lin. Vespertilio murinus. Lin.
Pecotus. Geoff. Vesp. auritus. Lin. (Plecotus)
Barbastella. Gray. Vespertilio barbastellus. Gmel.
Class IV. QUADRIPEDES.
Body horizontal, hairy; thumbs not separate on the hind
or fore-legs : toes clawed or hoofed: mammae rarely pectoral .

sorts: pelvis perfect.
Sub-class I. UNGUIGULATA. - Toes three, or five, clawed.
Ord. I. PTEROPHORAE
Teeth, cutting; canine, and grinders : mammae pectoral:
skin of the body extanded, uniting.the legs and tail, so as to
form a parachute: Toes, five, clawed: claws curved, acute:
coecum very large.
Fam. 1. GALEOPITHECIDAE - Teeth, two cutting above,
toothed ; six below, four middle ones cut, pectinated: canine
obtusely tubercuIar, short; like the grinders.
Galeopithecus. Geoff. Lemur volans. Lin.
____________________________________________________________________________
CHIROPTERA CLASSIFICATION - HIGH LEVEL

Essas divisões não são simplesmente arbitrárias e morfológicas...são representações da Evolução dos Morcegos...dos CHIROPTERA. Em cima um Pteropodidae, GRANDE com suas asas de Arcanjo, representando o centro de ORIGEM. No Centro, a América e o Velho Mundo. Irmãos muito semelhantes e derivados. Cherubins menores, nas escadarias do céu. Finalmente, o Grande Centro Evolucionário do Planeta: As famílias de menor tamanho a Evolução Pedomórfica Final.
Such divisions are not simply arbitrary and morphological...are representations of the Evolution of Bats ...the Chiroptera. On top a Pteropodidae, BIG with their wings of Archangel, representing the center of ORIGIN. In the Center, the America and the Old World. Very similar and derivatives brothers. Smaller Cherubins on the steps of heaven. Finally, the Biggest Evolutionary Center of the PLANET: The smaller families the Ultimate Paedomorphic Evolution.
De acordo com a evolução, como deve ficar a classificação dos Chiroptera ?
Ordem : Chiroptera
- subordem : MEGACHIROPTERA
superfamilia : Pteropodoidea
- subordem : MICROCHIROPTERA
-Infraordem : Noctiliformes
superfamilia : Rhinolophoidea
Familia: Megadermatidae - Megaderma, Macroderma, Lavia e Cardioderma
Familia: Nycteridae - Nycteris
Familia: Rhinolophidae - Anthops, Asellia, Aselliscus, Cloeotis, Coelops, Hipposideros, Paracoelops. Rhinonicteris, Rhinolophus, Paratriaenops e Triaenops.
superfamilia : Phyllostomoidea
superfamilia : Noctilionoidea
Familia: Noctilionidae - Noctilio
Familia: Mormoopidae - Mormoops e Pteronotus
Familia: Mystacinidae - Mystacina
-Infraordem : Vespertiliformes
superfamilia : Nataloidea
Familia: Myzopodidae - Myzopoda
Familia: Thyropteridae - Thyroptera
Familia: Natalidae - Natalus, Chilonatalus e Nyctiellus
Familia: Furipteridae - Furipterus e Amorphochilus
superfamilia : Vespertilionoidea
Familia: Myotidae - Myotis, Kerivoula, Murina
Familia: Plecotidae - Barbastella, Plecotus, Corynorhinus, Idionycteris e Euderma
Familia: Laephotidae - Laephotis
Familia: Otonycteridae - Otonycteris
Familia: Antrozoidae - Antrozous e Bauerus
Familia: Nyctophilidae - Nyctophilus e provavelmente Pharotis.
Familia: Craseonycteridae - Craseonycteris
Familia: Rhinopomatidae - Rhinopoma
O crânio dos Chiroptera

Chiroptera Evolution


PADRÕES BASAIS

Phyllostomidae Evolution I

Phyllostomidae Evolution II

PHYLLOSTOMIDAE EVOLUTION III

Phyllostomidae Evolution IV

Desmodus EVOLUTION

Desmodus EVOLUTION


Carollia Phylogeny (some relations) and sisters

Comparation - Phyllostomus-Carollia-Rhinophylla
 Evident paedomorphic sequence, to show that Rhinophylla is part of Carollia.
Evident paedomorphic sequence, to show that Rhinophylla is part of Carollia.
The progressive reduction of the molars shows a series of transformations.
The teeth are out of scale, just to compare series of transformations.
Rhinophylla is Carollia.

Stenodermatinae molars compared

Sturnira Evolution 1

Sturnira Evolution 2

Sturnira Biogeography

STENODERMATINA

NOCTILIONOIDEA (Skull and rhinarium)

Let us first deal with the most important question, concerning to the map, which I present here. How this map was built?
Dispersion of the Plecotidae, Antrozoidae and Histiotidae

Mapa Microchiroptera tropical

EVOLUTION in RHINOLOPHOIDEA

Rhinarium - Synapomorphic Nose Leaf

- subordem : MEGACHIROPTERA
superfamilia : Pteropodoidea
Familia: Pteropodidae - Macroglossus, Megaloglossus, Eonycteris, Syconycteris, Melonycteris, Notopteris, Eidolon, Rousettus, Boneia, Myonycteris, Pteropus, Acerodon, Neopteryx, Pteralopex, Styloctenium, Dobsonia, Aproteles, Harpyionycteris, Plerotes, Hypsignathus, Epomops, Epomophorus, Micropteropus, Nanonycteris, Scotonycteris, Casinycteris, Cynopterus, Megaerops, Ptenochirus, Dyacopterus, Chironax, Thoopterus, Sphaerias, Balionycteris, Aethalops, Penthetor, Haplonycteris, Otopteropus, Alionycteris, Latidens, Nyctimene, Paranyctimene, Mirimiri
- subordem : MICROCHIROPTERA
-Infraordem : Noctiliformes
superfamilia : Rhinolophoidea
Familia: Megadermatidae - Megaderma, Macroderma, Lavia e Cardioderma
Familia: Nycteridae - Nycteris
Familia: Rhinolophidae - Anthops, Asellia, Aselliscus, Cloeotis, Coelops, Hipposideros, Paracoelops. Rhinonicteris, Rhinolophus, Paratriaenops e Triaenops.
superfamilia : Phyllostomoidea
Familia: Phyllostomidae - Chrotopterus, Glyphonycteris, Lampronycteris, Lonchorhina, Lophostoma, Macrophyllum, Macrotus, Micronycteris, Mimon, Phylloderma, Phyllostomus, Tonatia, Trachops, Trinycteris, Vampyrum, Artibeus, Ametrida, Ardops , Ariteus, Centurio, Chiroderma, Ectophylla, Enchisthenes, Mesophylla, Phyllops, Platyrrhinus, Pygoderma, Sphaeronycteris, Stenoderma, Uroderma, Vampyressa, Vampyrodes, Vampyrops, Sturnira, Desmodus, Diaemus, Diphylla, Brachyphylla, Erophylla, Phyllonycteris, Anoura, Choeroniscus, Choeronycteris, Glossophaga, Hylonycteris, Leptonycteris , Lichonycteris, Monophyllus, Musonycteris, Scleronycteris, Lionycteris, Lonchophylla, Platalina, Xeronycteris e Carollia.
superfamilia : Noctilionoidea
Familia: Noctilionidae - Noctilio
Familia: Mormoopidae - Mormoops e Pteronotus
Familia: Mystacinidae - Mystacina
-Infraordem : Vespertiliformes
superfamilia : Nataloidea
Familia: Myzopodidae - Myzopoda
Familia: Thyropteridae - Thyroptera
Familia: Natalidae - Natalus, Chilonatalus e Nyctiellus
Familia: Furipteridae - Furipterus e Amorphochilus
superfamilia : Vespertilionoidea
Familia: Myotidae - Myotis, Kerivoula, Murina
Familia: Plecotidae - Barbastella, Plecotus, Corynorhinus, Idionycteris e Euderma
Familia: Vespertilionidae - Arielulus, Eptesicus, Hesperoptenus, Dasypterus, Lasiurus, Nycticeinops, Nycticeius, Rhogeessa, Scoteanax, Scotoecus, Scotomanes, Scotophilus, Scotorepens, Glischropus, Nyctalus, Parastrellus, Perimyotis, Pipistrellus, Scotozous, Chalinolobus, Eudiscopus, Falsistrellus, Glauconycteris, Niumbaha, Hypsugo, Ia, Mimetillus, Neoromicia, Philetor, Tylonycteris, Tomopeas, Vespadelus e Vespertilio.
Familia: Histiotidae - HistiotusFamilia: Laephotidae - Laephotis
Familia: Otonycteridae - Otonycteris
Familia: Antrozoidae - Antrozous e Bauerus
Familia: Nyctophilidae - Nyctophilus e provavelmente Pharotis.
Familia: Craseonycteridae - Craseonycteris
Familia: Rhinopomatidae - Rhinopoma
Familia: Molossidae - Mormopterus, Sauromys, Platymops, Molossops, Cynomops, Myopterus, Tadarida, Chaerephon, Mops, Otomops, Nyctinomops, Eumops, Promops, Molossus, Cheiromeles
Familia: Emballonuridae - Saccolaimus, Taphozous, Mosia, Emballonura, Coleura, Cormura, Saccopteryx, Rhynchonycteris, Centronycteris, Peropteryx, Balantiopteryx, Cyttarops, Diclidurus.
Vejam algumas das características na evolução do crânio dos Chiroptera.
Os Pteropodidae são os mais conservadores: os processos pós-orbitais são desenvolvidos, a mandíbula tem um processo angular largo e arredondado, o crânio é curvo dorsalmente, as órbitas são grandes e um pouco voltadas para frente, todas essas características são adaptações à VIDA arborícola.
 |
| A EVOLUÇÃO DO ENCÉFALO DOS CHIROPTERA - Nos Microchiroptera a redução do focinho e do olho, levou a uma redução do telencéfalo, deixando o cerebelo proporcionalmente maior. A polarização dos caracteres cranianos depende desse entendimento: a redução por pedomorfose deixou os crânios semelhantes aos dos jovens, focinhos curtos com caixas cranianas grandes. Na figura temos a condição plesiomórfica encontrada na linha de base dos Phyllostomidae e dos Vespertilionidae. Partindo disso, iremos entender a evolução dos encéfalos dos Chiroptera. |
Nos Microchiroptera o crânio sofreu uma redução de tamanho, algumas características mudaram e outras foram perdidas. Phyllostomus é intermediário entre os megabats e microbats. Está na base da evolução desse grupo.
Chiroptera Evolution
 |
| Na figura acima vemos crânios em vistas ventrais e encéfalos em vistas dorsais (em cores) |
Estou apresentando apenas alguns grupos estrategicamente para simplificar o entendimento. Veja também as 2 figuras superiores.
EVOLUÇÃO DOS CHIROPTERA e a DIVERSIFICAÇÃO DOS CRÂNIOS
![O quadro acima foi construído para mostrar as diferenças de forma e tamanho que existem nos CHIROPTERA. Dessa maneira podemos VER a EVOLUÇÃO! Na linha de baixo vemos os morcegos de maior porte, os Pteropodidae e os Microchiroptera basais, exclusivamente tropicais : Phyllostomoidea e Rhinolophoidea. Ainda temos os Myotidae [Myotinae e Murinae] que representam uma transição entre os de maior tamanho e os pequenos morcegos, que se encontram acima. A área de origem dos Rhinolophidae se situa entre a África e a Índia, região que chamei de "região de quebra das florestas".](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEjOTIT7fuZw8o7olzhaHZOJmaGdtgSMjrwhbf_TDRoKqc1e3zTurEoKtdDHQsfkxTgSyp54dh2pQ2zh_F9NRKDUzoERbBXzXZuxB2MB-rV7kQeQTPJTzD-BZ0S8UFdfqyryPUjH2QIEopMOPRctgkvvPPRexU9ww_8Ybd6u7MB9bIuWtgPbt9P7lT2xKA/s5291/03.jpg) |
| O quadro acima foi construído para mostrar as diferenças de forma e tamanho que existem nos CHIROPTERA. Dessa maneira podemos VER a EVOLUÇÃO! Na linha de baixo vemos os morcegos de maior porte, os Pteropodidae e os Microchiroptera basais, exclusivamente tropicais : Phyllostomoidea e Rhinolophoidea. Ainda temos os Myotidae [Myotinae e Murinae] que representam uma transição entre os de maior tamanho e os pequenos morcegos, que se encontram acima. A área de origem dos Rhinolophidae se situa entre a África e a Índia, região que chamei de "região de quebra das florestas". |
O quadro acima permite também percebemos que os Pteropodidae por serem mais próximos à base da evolução dos Chiroptera e estarem ligados primitivamente as áreas de origem, retiveram um tamanho maior que os demais grupos e os Phyllostomoidea e Rhinolophoidea divergiram muito pouco desse modelo.
A minha interpretação desses fatos é semelhante àquela que já vimos anteriormente para as regiões biogeográficas da América do Sul. Os primeiros morcegos que surgiram, eram do porte dos Pteropodidae, mas tinham dentes como os Phyllostomidae. Eles se dispersaram para o oeste junto com a expansão das florestas, em uma fase úmida da história climática do planeta. De acordo com os registros fósseis conhecidos, isso se deu por volta do inicio do Cenozóico. Como nessa época os continentes já estavam com seus contornos definidos, ou seja, como são hoje, a dispersão se deu seguindo pela Índia até a África central. Provavelmente nessa época ainda formavam um grupo único e homogêneo, como uma espécie em expansão.
A África nessa época, de acordo com os dados inferidos através dos sedimentos da crosta, já havia se quebrado da América do Sul, mas ainda deveria estar muito próxima. A dispersão para a América do Sul, mais provavelmente ocorreu através do mar. Nesse momento a população primitiva dos morcegos que originariam os Phyllostomidae teria colonizado a América do Sul.
Com o distanciamento da África e a abertura do Oceano Atlântico, o clima da África deve ter se alterado drasticamente. Isso realmente parece estar evidente nas transformações que podemos observar nos Megadermatidae. Se compararmos Vampyrum e Phyllostomus que são os mais conservadores morcegos do Novo Mundo, com Lavia e Cardioderma [África], Megaderma [Índia até Indonésia] e Macroderma [Austrália], constatamos que existe um aumento de tamanho em direção à Austrália. Esses morcegos são todos muito semelhantes, mas Lavia e Cardioderma são menores e um pouco mais diferenciados. Interpreto isso, como uma transformação maior do ambiente na África, com empobrecimento da vegetação, gerando diminuição no tamanho. [veja Raven, P. H. & Axelrod, D. I. 1974. Angiosperm Biogeography and Past Continental Movements.]
Depois da fragmentação basal surge o ramo que deu origem aos demais grupos. Na base desses grupos menores está Myotidae. Podemos dizer que Myotidae se destacou dos demais, no mesmo lugar de quebra climática que se originaram posteriormente os Rhinolophidae, entre a África e a Índia, nessa região que hoje é desértica.
Ali mesmo, por influência da diagonal seca da Ásia, sempre se estabelece essa quebra entre as duas grandes florestas do Velho Mundo. Sendo portanto uma importante área de diferenciação.
Myotidae tem no gênero Myotis seu representante mais significativo, tanto pela diversificação de espécies, quanto pela sua ampla distribuição. Pode-se dizer que enquanto o gênero Myotis se diversificou, todos os outros grupos de Microchiroptera também o fizeram. Observando os grupos que estão acima dessa linha inferior, temos Plecotidae contra o resto. Plecotidae é um grupo que ainda retém as grandes orelhas, mas já apresenta uma diminuição de tamanho e encurtamento do focinho, que caracteriza todos os outros. O grupo-irmão de Plecotidae se caracteriza por um achatamento craniano que observamos em Vespertilionidae e em todos os demais.
Vespertilionidae se caracteriza por um aumento progressivo de tamanho, culminando em Scotophilus e Lasiurus, e seu grupo-irmão origina Emballonuridae que também vai apresentar um aumento inicial no tamanho, seguido de nova diminuição.
O próximo grupo a surgir é formado por Rhinopomatidae e Molossidae, que junto com Emballonuridae, formam um grupo de morcegos com a cauda livre e orelhas mais reduzidas de tamanho e com várias transformações como redução do tragus e avanço ao redor dos olhos. A dentição é derivada com amplas bacias hipoconais nos molares superiores.
Observem as cristas associadas à região do cerebelo e bulbo olfativo. A redução do telencéfalo está associada à redução da crista sagital. Em Molossidae e Rhinopomatidae a grande redução do telencéfalo produziu uma crista secundária, formada pela região do cerebelo e da região do lóbulo olfativo. Essas idéias são quase todas inéditas.
__________________________________________________________________________________________
MEGACHIROPTERA


Quando tentei entender a diversidade e complexidade dos megachirópteros, me deparei com duas linhas evolutivas, uma africana e outra asiática. Entre elas os grandes morcegos que com suas grandes asas colonizaram metade da superfície do planeta, indo até as ilhas mais remotas e marcando sua presença desde Madagáscar até Tonga e além....falo exatamente de Pteropus.
Mas o destino dessas formas de vida, parece ser outro: a polinização, pois tanto na linha africana como na asiática a tendência evolutiva foi a miniaturização, culminando num grupo polinizador especializado. A vida tem sido sempre uma resposta ao preenchimento do espaço deixado pela devastação e destruição que o tempo inevitavelmente trás.
Acredito que essa miniaturização culminou com o aparecimento dos microchiroptera. Pois minha visão da Deriva Continental me leva a pensar que grandes morcegos chegaram até a América quando esta ainda estava acessível para o voo de um "pteropodídeo".
Mas aí inevitavelmente a tendência polinizadora fez a sua parte...pois voar é ligar todas as partes...é conectar a vida das plantas....que é base de toda vida.
Mas quando o tempo também fez sua parte na renovação...esse Shiva impiedoso....restaram os pequenos polinizadores e insetívoros que estão sempre na base de populações de ilhas, refazendo a Natureza e recompondo esse quadro eterno.
Na Austrália os pequenos marsupiais tiveram um importante papel na polinização e renovação da vida depois do Cretáceo....pois as flores estavam lá esperando seus admiradores....elas também precisavam deles.
Antes do aparecimento das plantas com flores e frutos muitas das formas de vida que conhecemos hoje nem tinham razão de ser.
A América ficou isolada do resto do mundo...no norte o contato ainda existia através de Bering...mas no sul apenas uma ilha devastada exposta a influência das correntes marítimas do Pacífico. Só com o aparecimento dos Andes é que a umidade pode ser retida, principalmente na Amazônia para manter uma grande diversidade...pois as florestas do leste...filhas das chuvas orográficas, que existem em todos os continentes, não comportam uma grande massa.
Observamos que a Australia é mais ou menos isso...o que foi a América do Sul....lá está tudo preservado nas florestas do leste....muitas relíquias como as minúsculas populações de Burramys que se isolaram num último refúgio nas frias montanhas do sul.
Os [Cynopterinae + Macroglossinae + Latidens] formam um grupo monofilético.
Nyctimene e Paranyctimene devem ser incluidos dentro dos Cynopterinae, de novo a velha tradição gradista...que separa aquilo que é bem derivado.
Os gêneros Notopteris, Eonycteris, Melonycteris, Macroglossus e Syconycteris, são adaptados para uma dieta a base de polem e néctar. São morcegos polinizadores. Sua evolução parece ser independente da evolução dos morcegos polinizadores africanos. Talvez representem o grupo monofilético mais coerente de todos os Pteropodidae asiáticos.
MICROCHIROPTERA
Entre os Microchiroptera, Vampyrum e Phyllostomus apresentam crânios e encéfalos mais conservadores, por isso são aqui considerados como basais, o crânio de Vampyrum é muito conservador, assim como o encéfalo, porém temos um palato estreito [uma derivação talvez associada a carnivoria].
Certamente ter uma folha nasal não pode ser considerado um caráter basal, mas o tamanho e a forma do crânio e o encéfalo, SIM.
Os Vespertilionidae e os Emballonuridae não possuem folha nasal isso é PLESIO, certamente ! Mas existem várias outras característica que são modificações.
Como um focinho bem curto e a região das pré-maxilas muito modificada. O corpo de modo geral é muito pequeno. Todas essas características são derivadas. O encéfalo particularmente é bem pedomórfico !
Temos então uma heterobatmia bem nítida nesse contexto basal.
Em cada um desses grupos existe pelo menos uma modificação estrutural bem importante. Ter uma folha nasal não significa simplesmente "MORFOLOGIA", essas estruturas estão muito provavelmente associadas a essas orelhas grandes, como um mecanismo de pergunta e resposta. Penso em uma forma de comunicação entre os membros da mesma espécie !
Pelo outro lado a modificação da região das pré-maxilas e a importante diminuição de tamanho representou um avanço ecológico surpreendente ! As orelhas em alguns Vespertillionidae também são significativas, mas a relação é outra, a comunicação entre os membros da mesma espécie deve ter sido o primeiro passo evolutivo. O uso na alimentação deve ter sido uma evolução posterior. A modificação das pré-maxilas é comum a vários grupos de morcegos, nos basais Pteropodidae, são pouco modificadas, assim como nos Phyllostomidae, esses vários passos no sentido de modificar essa região representam uma adaptação importantíssima....não é simplesmente morfologia.
O TEMPO da PALAVRA "MORFOLOGIA" há muito já terminou para mim.
Penso em forma-função sempre......afinal....pra que estudar morfologia ?

The Basal Patterns that will guide us in the final understanding the characters of Phyllostomoidea-Rhinolophoidea.
Note the conservative nasal leaf of Vampyrum and Chrotopterus ...it is slightly modified in Macroderma. Also note the reduction of the upper lip in Phyllostomus and even more in Phylloderma...the size and shape of the ears ... reduced in Phyllostomus and Phylloderma. The shortening of the snout, more pronounced in Phyllostomus.
The modification of the lower lip in Phyllostomus and Phylloderma.
All these bats have relatively large eyes ... so the eye reduction that is observed in the smaller Phyllostomidae bats, or in Nycteridae and Rhinolophidae are derived characters.
Also, the reduction or loss of premolars that usually accompanies the shortening of the muzzle must be regarded as derivative, both in Rhinolophoidea as in Phyllostomoidea.
The shortening of the palate also usually related to loss of molars, usually the third molar, must be regarded as derivative.
See this FIGURE on biggest Increase to appreciate the details ! Spectacular !
Apresento aqui os Padrões Basais que nos guiarão na compreensão definitiva dos caracteres dos Phyllostomoidea-Rhinolophoidea.
Observe a folha nasal conservadora em Vampyrum e Chrotopterus...está um pouco modificada em Macroderma.
Observe a redução do lábio superior em Phyllostomus e mais ainda em Phylloderma.
Observe o tamanho e forma das orelhas...reduzidas em Phyllostomus e Phylloderma.
O encurtamento do focinho mais pronunciado em Phyllostomus.
A modificação do lábio inferior em Phyllostomus e Phylloderma.
Todos esses morcegos têm olhos relativamente grandes...portanto a redução dos olhos que se observa nos morcegos Phyllostomidae menores, ou em Nycteridae e Rhinolophidae, são caracteres derivados.
Também a redução ou a perda de pré-molares que geralmente acompanha o encurtamento do focinho deve ser considerada como uma derivação, tanto nos Rhinolophoidea como nos Phyllostomoidea.
O encurtamento do palato também costuma estar relacionado à perda de molares, geralmente o terceiro molar.

Limites da pedomorfose - Um morcego nunca terá o tamanho de uma formiga. Existe um limite de tamanho para cada organismo....um mínimo no números de células.
O processo pedomorfico mostrado pelos CHIROPTERA, nos dá uma ideia desse limite de tamanho mínimo.
Evolução e variação populacional - especiação gradual
O processo de especiação pode ocorrer de maneiras diferentes, dependendo de fatores ambientais como a evolução de biomas ou a degradação desses mesmos biomas (que também não deixa de ser uma evolução). Pode também depender de processos ocupacionais de determinadas áreas (ambientes) ou de fragmentação desses ambientes. O isolamento geográfico pode fragmentar biomas inteiros em unidades menores, como também submeter populações inicialmente semelhantes a diferentes níveis de degradação, gerados por gradientes climáticos ou ilhas resultantes da fragmentação desses gradientes.
Em um único processo de especiação podem ser geradas inúmeras espécies, que são o resultado de fragmentações de gradientes climáticos.
Os gêneros Vampyrum e Phyllostomus estão associados às bases dos dois grandes ramos evolutivos da familia Phyllostomidae. Esses gêneros apresentam tamanho grande, são dois remanescentes pouco modificados das duas populações basais que deram origem à família. O ramo de Vampyrum deu origem aos morcegos de crânio longo que culminaram nos POLINIZADORES e o ramo de Phyllostomus deu origem aos morcegos FRUGÍVOROS. Os demais gêneros, aparentemente, se originaram de um gradiente intermediário que existiu entre Vampyrum e Phyllostomus. Esse gradiente se fragmentou, dando origem à morcegos proximamente ligados a Vampyrum e Phyllostomus, gerando uma distribuição de caracteres em mosaico.
O gênero Mimon está no ramo de Vampyrum, mas compartilha caracteres derivados com Macrophyllum, Lonchorhina e Phylloderma, que pertencem ao ramo de Phyllostomus, mas por sua vez conservam caracteres plesiomórficos basais. Polinizadores especializados surgiram independentemente nesses dois grandes grupos basais.
Limits of paedomorphosis - A bat will never have the size of an ant. Is there a size limit for each organism .... at least in cell numbers.
The paedomorphic process shown by CHIROPTERA, gives us an idea of that minimum size limit.
Evolution and populational variation - gradual speciation
The process of speciation can occur in different ways, depending on environmental factors such as the evolution of biomes or degradation of those biomes. May also depend on occupational processes of certain areas (environments) or fragmentation of these environments. Geographic isolation can fragment entire biomes into smaller units, as well as submit initially similar populations at different levels of degradation, generated by climatic gradients and islands resulting from the fragmentation of these gradients.
In a unique process of speciation, many species may be generated, which are the result of fragmentation of climatic gradients.
Phyllostomidae Evolution II

Apresento aqui a evolução dos Phyllostomidae na forma de um cladograma....mesmo sabendo que um cladograma nunca poderia ser considerado como a melhor maneira de representar as relações filogenéticas de um grupo....pois "as radicais dicotomias" não expressam bem as distribuições em mosaico observadas no conjunto dos caracteres.
I present here ... the evolution of Phyllostomidae as a cladogram..... even knowing that one cladogram could never be considered as the best way to represent the phylogenetic relationships of a group .... because "the radical dichotomies" do not express well the mosaic distributions observed in the set of characters..

The Origin of the PHYLLOSTOMIDAE bats.
The genera Vampyrum and Phyllostomus are associated with the bases of the two major evolutionary branches of the family Phyllostomidae.
These genres have great size, are two remaining little changed, of the two baseline populations that gave rise to the family.
The Vampyrum branch gave rise the bats with long skull, which culminated in POLLINATORS and Phyllostomus branch gave rise to frugivorous bats.
The other genera apparently originated from an intermediate gradient that existed between Vampyrum and Phyllostomus. That gradient fragmented, giving rise to bats closely linked to Vampyrum and Phyllostomus, generating a mosaic distribution of characters.
The genus Mimon is in the branch of Vampyrum and Chrotopterus, but shares derivatives characters with Macrophyllum, Lonchorhina and Phylloderma that belong to the branch of Phyllostomus, these genres in turn retain basal plesiomorphic characters, like 3 premolars in the mandible. Phyllostomus and the frugivorous bats have lost one premolar in each half mandible, having short muzzles. But specialized pollinators bats have arose independently in the two large basal branchs.
Mimon, Macrophyllum and Lonchorhina share derived characters such as very small eyes, nasal leaves proportionally long compared to their heads, large pointed ears, short muzzles and small sized bodies. These characters are true synapomorphies in a MOSAIC distribution.
However an important ecological BREAK should have defined two intermediate branches.
One of these branches formed by Phylloderma, Desmodus, Lonchorhina and Macrophyllum, that retained 3 premolars in each half mandible (with a central premolar much reduced, in a shortened mandible) and the other branch formed by Mimon (with a relatively elongated mandible as well as the palate, and only 2 plesiomorphic premolars in each half mandible).
It seems to have been a "CONTINUUM" between all these genres. For this reason I termed gradual ecological speciation. As they all have wide distribution and dominant species with distribution sometimes covering the whole warmer range of the Neotropical Region, they must have originated from a highly variable species, which divided following ecological paths.
The speciation of the genres mentioned above can not be explained by geographic isolation, as in the case of more recent species. However the miniaturizations of both Carollia or Micronycteris seem to be associated with isolation in Central America as demonstrated in the table "Evolution of Carollia-Phyllostomus and Sturnira-Brachyphylla".
Veja a próxima figura para melhor compreensão:
LOOK this picture in biggest SIZE for details !
Parece ter existido um CONTINUUM entre todos esses gêneros. Por isso denominei especiação ecológica gradual. Como todos eles possuem ampla distribuição e espécies dominantes com distribuição as vezes abrangendo toda faixa mais quente da Região Neotropical, devem ter se originado a partir de uma espécie altamente variável, que se dividiu seguindo rumos ecológicos.
A especiação desses gêneros citados acima não pode ser explicada por isolamentos geográficos, como no caso das espécies mais recentes. No entanto as miniaturizações tanto de Carollia como de Micronycteris parecem estar associadas a um isolamento na América Central como demonstrei na prancha "Evolução de Phyllostomus-Carollia e Sturnira-Brachyphylla".
Phyllostomidae Evolution IV

The two basal forms Vampyrum and Phyllostomus derived each for a different pole.
The genus Vampyrum and their descendants were characterized by an elongation of the snout, with long and narrow palates, and with mandibular canines very close. Vampyrum is an extreme, has a long and distinguished mandibular symphysis and diastemas between the upper premolars.
The genus Phyllostomus and their descendants have lost a premolar in each half mandible (p3), resulting in shorter snouts, with wider palates. The Macrophyllinae, their immediate sister group, still retains the middle premolar (p3) greatly reduced.
Between these two early trends developed a gradient of ecological forms.
The phylogenetic pattern of Phyllostomidae is very peculiar, it suggests that an ancestral species arrived in a big new continent, occupying a large space, a large community under development. This species has diversified but maintained for a long time their genetic links.
Os gêneros Vampyrum e Phyllostomus são os maiores em tamanho, entre os filostomídeos. Possuem várias características basais já comentadas no quadro "Padrões BASAIS". Eles são conservadores em vários aspectos, e mesmo possuindo suas inevitáveis derivações e suas autapomorfias, podem ser considerados antigos remanescentes das duas populações basais que deram origem à famíla.
A ampla análise que realizei na família Phyllostomidae, mostrou que uma população ancestral vinda da África colonizou a América do Sul, espalhando-se provavelmente por toda região tropical. Essa espécie deve ter se polarizado em duas tendências ecológicas. Uma delas derivou para insetivoria-carnivoria (linha de Vampyrum) e outra para insetivoria-néctar e pólen (linha de Phyllostomus). Por serem animais voadores, com um potencial de dispersão muito maior que qualquer outro tipo de mamífero, sua evolução não se compara a de nenhum outro grupo e sua diversificação obedeceu a regras totalmente diferentes.
A análise do cladograma básico e as comparações dos caracteres, mostraram duas grandes populações iniciais, com um gradiente entre elas, que se quebrou deixando padrões em mosaico entre as espécies menores no centro desse gradiente.
O centro dessa quebra ocorreu entre os gêneros Mimon e Macrophyllum. Esses gêneros compartilham características derivadas, surgidas durante a fase que antecedeu a "grande quebra ecológica". Eu estou falando da quebra de um gradiente contínuo que existiu entre as duas populações basais de morcegos conservadores Vampyrum e Phyllostomus.
A população inicial vinda da África deve ter desenvolvido uma grande variação de formas ecológicas, que por um longo tempo, mantiveram um gradiente de formas intermediárias. Devido a grande capacidade de dispersão desses mamíferos, nem podemos pensar em isolamento reprodutivo. Sem dúvida, sua evolução foi uma evolução ecológica, como mostra a análise da distribuição dos caracteres. Por esse motivo nem a "mentalidade gradista", nem a "cladística" conseguiram resolver as intrincadas distribuições em mosaico desses caracteres.
A "mentalidade gradista" é primitiva e grosseira, é natural e trabalha da mesma forma em todos os grupos animais. Separa sempre por semelhanças e diferenças. Sempre detecta em primeiro lugar as tendências marcantes, como "morcegos polinizadores de crânios longos" e "morcegos frugívoros de crânios curtos", e por isso sempre cria um terceiro grupo parafilético (um saco de gatos), ou seja, "O RESTOLHO" !
As duas formas basais, Vampyrum e Phyllostomus derivaram cada uma para um polo diferente.
O gênero Vampyrum e seus descendentes se caracterizaram por um alongamento do focinho, com palatos longos e estreitos, e com caninos mandibulares muito próximos. Vampyrum é um extremo, possui uma longa e distinta sínfise mandibular e diástemas entre os pré-molares superiores.
O gênero Phyllostomus e seus descendentes perderam um pré-molar em cada metade da mandíbula (p3), resultando em focinhos mais curtos, com palatos mais largos. Os Macrophyllinae seu grupo irmão imediato, ainda retem o pré-molar do meio (p3) bem reduzido.
Entre essas duas tendências iniciais desenvolveu-se um gradiente de formas ecológicas. O padrão filogenético dos Phyllostomidae é muito peculiar, ele sugere que uma espécie ancestral, chegou em um novo e grande continente, ocupando um grande espaço, uma grande comunidade em desenvolvimento.
Essa espécie se diversificou mas manteve por longo tempo seus vínculos genéticos.

A compreensão das relações filogenéticas dos morcegos hematófagos dentro da família Phyllostomidae pode ser considerada como o maior desafio de todos nessa empreitada que venho desenvolvendo.
O crânio desses morcegos é provavelmente o mais afetado pelo processo pedomórfico, se comparado aos demais membros desse grande grupo. Eles possuem um crânio ovalado, com arcos zigomáticos muito pouco expandidos lateralmente, número de dentes muito reduzido, e muito especializados, praticamente sem molares. Essa simplificação do crânio se reflete exteriormente na redução das orelhas, na redução do comprimento do focinho, simplificação da folha nasal e redução do lábio superior.
Dois pontos importantes foram levados em consideração para se detectar suas relações filogenéticas: 1) Esses morcegos possuem um crânio muito encurtado, isso sugere uma afinidade com o ramo evolutivo de Phyllostomus, já que o ramo de Vampyrum se destaca por uma evolução caracterizada por um alongamento da mandíbula e do palato. 2) Eles por sua vez conservam a condição basal de 3 pré-molares na mandíbula, enquanto no clado de Phyllostomus houve perda de um dente (p3).
Phylloderma é um gênero com um crânio relativamente ovalado, sendo até globoso nos indivíduos mais jovens. Assim como os hematófagos, também possui arcos zigomáticos pouco expandidos lateralmente e focinho relativamente curto, se comparado ao clado de Phyllostomus. Possuem 3 pares de pré-molares na mandíbula, com redução de tamanho em p3. Essa condição já aponta o rumo da perda de p3 (condição obrigatória), que aconteceu no clado de Phyllostomus.
Macrophyllum e Lonchorhina são 2 gêneros que também possuem focinho curto com presença de 3 pré-molares na mandíbula com redução em p3. Esses gêneros compartilham também outra condição derivada, redução dos pré-molares das maxilas, P3 é bem menor que P4 nesses dois gêneros. Os morcegos hematófagos possuem apenas 1 par de pré-molares superiores, provavelmente P4 (aparentemente a redução culminou em uma perda de P3). Esses dois gêneros já foram considerados como gêneros afins por vários autores, pois além dessas características cranianas compartilham outros caracteres derivados, como olhos muito pequenos, focinho curto, grandes orelhas pontudas (intermediárias entre as grandes e arredondadas orelhas basais do ramo de Vampyrum e as orelhas mais curtas e pontudas do clado de Phyllostomus. Suas longas folhas nasais, quando comparadas ao tamanho de suas cabeças, são as mais desenvolvidas de todos os Phyllostomidae.
Os hematófagos contrastam grandemente com Macrophyllum e Lonchorhina por possuírem folhas nasais reduzidas, orelhas reduzidas, olhos relativamente grandes e maior tamanho corpóreo. Mas são intermediários entre Phylloderma e esses dois gêneros em tamanho e nas características dos pré-molares. Somando-se a isso Desmodus e Lonchorhina compartilham os hábitos comportamentais citados acima, só deixam seus abrigos após a total escuridão, diferentemente de outros morcegos. Também foram detectados flagelados tripanossomatídeos em Lonchorhina (estritamente insetívoro), ainda que numa frequência comparativamente baixa, se comparada à Desmodus. Esses dois pontos acabaram contribuindo de uma forma modesta para se elaborar uma hipótese sobre a origem da sanguivoria nos Phyllostomidae, pois somados às outras características já citadas, nos apontam uma direção. Alguma coisa realmente palpável num mar de suposições sem fundamento, como é o quadro externo.
Chama atenção também a semelhança morfológica entre as orelhas de Desmodus e Lonchorhina, por possuírem um contorno especialmente diferente. O padrão apresentado por Desmodus poderia ser considerado uma redução a partir de um tipo como as de Lonchorhina. Também os lábios inferiores desses morcegos se assemelham, apesar de serem praticamente idênticos aos mais basais. No entanto poderiam ser considerados derivados, pois Macrophyllum e Phylloderma (da mesma subfamilia) possuem verrugas no lábio inferior, assim como Mimon (na base do ramo de Vampyrum) e praticamente todo o clado de Phyllostomus (grupo irmão imediato dos Macrophyllinae).
Observação : flagelados tripanossomatídeos já foram encontrados em muitas espécies diferentes de morcegos. Em várias espécies de Artibeus, em Sturnira, Carollia, Glossophaga, Molossus, etc.
Como ocorreu essa infestação nesses morcegos ? Carrapatos ?
E porque isso não acontece em certas regiões ?
A subfamilia MACROPHYLLINAE aqui proposta, é composta pelos gêneros Phylloderma, Diphylla, Desmodus, Diaemus, Lonchorhina e Macrophyllum, tem como característica principal a manutenção de 3 pares de pré-molares na mandíbula, p2, p3 e p4, com p3 mais reduzido que os demais, comprimido entre p2 e p4.
Os gêneros Diphylla, Desmodus e Diaemus formam um subgrupo monofilético de morcegos hematófagos, com incisivos altamente especializados.
Essa subfamília apresenta alguns caracteres com distribuição em mosaico e tem como grupo irmão imediato o clado formado por Phyllostomus, Carollia, Sturnira, Brachyphylla, Erophylla, Phyllonycteris + os Sternodermatinae.
Se quisermos manter as tradicionais subfamílias, então a classificação ficaria assim:
Familia PHYLLOSTOMIDAE (Phyllostomus subgroup)
Subfamilia : Macrophyllinae
Subfamilia : Phyllostominae
Subfamilia : Carolliinae
Subfamilia : Sturnirinae
Subfamilia : Brachyphyllinae
Subfamilia : Sternodermatinae

Aqui nesta figura vemos uma comparação das mandíbulas dos 3 gêneros de filostomídeos hematófagos, Diphylla, Desmodus e Diaemus. Diphylla possui 4 dentes pós-caninos enquanto Desmodus e Diaemus apenas 3.
No lado esquerdo da figura apresento duas sinapomorfias importantes, que apontam uma relação filogenética mais estreita entre os gêneros Desmodus e Diaemus.
1 - A perda do primeiro molar (m1), veja a comparação de Diphylla com Phylloderma (acima) e Desmodus (abaixo).
2 - Uma modificação dos pré-molares p2 e p3. Diphylla conservou a forma típica dos pré-molares, veja a comparação na parte central desse quadro com Phylloderma e Brachyphylla. O pré-molar inferior 3 (p3) está bem reduzido nos Macrophyllinae (Phylloderma, Lonchophylla, Macrophyllum mais os hematófagos) e ausente em Brachyphylla, assim como nos demais gêneros do clado de Phyllostomus (veja a comparação com Sturnira, Uroderma e Erophylla). Em Desmodus e Diaemus p2 e p3 apresentam uma inclinação para a região posterior, a cúspide central se deslocou para parte distal do dente. Assim como em Phylloderma p2 é maior que p3, mas apresenta uma borda laminar curva (caráter exclusivo de Desmodus e Diaemus).
Uma terceira sinapomorfia muito especial, ou seja um caráter "derivado" e compartilhado por Diaemus e Desmodus.
3 - Esses dois gêneros possuem um DIÁSTEMA entre os incisivos. Diphylla possui os 4 incisivos inferiores alinhados, existe um pequeno diástema junto aos caninos, "mas não entre os incisivos".
Desmodus e Diaemus possuem um diástema comparável ao de certos morcegos polinizadores. Um diástema entre os incisivos facilita sem dúvida os movimentos repetitivos da língua durante à alimentação baseada em líquidos.
Certamente essa característica deve refletir diferenças na tomada do alimento. Diphylla tem uma forte preferência pelo sangue de aves, enquanto Diaemus e Desmodus preferem o sangue de mamíferos.
Cada um desses gêneros retém caracteres plesiomórficos (mais conservadores) em comparação com os outros dois.
Diphylla possui os incisivos alinhados, enquanto Desmodus e Diaemus derivaram para um diástema entre os incisivos. Diphylla tem 1 molar em cada metade da mandíbula, os outros perderam.
Desmodus tem orelhas mais conservadoras, enquanto Diphylla e Diaemus possuem orelhas reduzidas, mais curtas e modificadas em vários aspectos.
Diaemus tem o processo coronóide da mandíbula mais plesiomórfico (semelhante ao de Phylloderma). Os outros dois são mais modificados cada um a sua maneira. Em Desmodus o côndilo mandibular é mais alto que o processo coronóide, sem dúvida o mais "fetal" de todos, um caráter derivado exclusivo (uma autapomorfia).
A comparação com Brachyphylla é importante, pelo fato desse gênero apresentar uma redução da folha nasal comparável à dos filostomídeos hematófagos (Diphylla, Desmodus e Diaemus). Brachyphylla apresenta também uma redução das orelhas e do focinho, resultantes de um processo pedomórfico evolutivamente independente. Veja no quadro seguinte deste álbum, que o lábio superior de Brachyphylla é muito semelhante aos dos morcegos hematófagos, quase um "lábio leporino". A pedomorfose afeta a base da folha nasal e o lábio superior de uma maneira semelhante em morcegos de diferentes clados. Como podemos ver nestes exemplos, os morcegos do clado de Phyllostomus perderam o pré-molar inferior 3 (p3), como resultado de um encurtamento do focinho ocorrido na base desse clado.
Observe que Phyllostomus tem cúspides pontudas nos molares da mandíbula, mas não tão evidentes como em Lonchorhina que é estritamente insetívoro. Phyllostomus se alimenta de pólen, néctar e complementa sua dieta com insetos. Brachyphylla e Erophylla se alimentam de pólen, néctar, frutas e insetos. Sturnira é frugívoro e Uroderma é primariamente frugívoro mas complementa sua dieta com insetos, pólen e néctar. Nesses 4 últimos gêneros os molares possuem cúspides mais baixas.
Em Erophylla (um polinizador especializado, Brachyphyllinae) houve um alongamento posterior do focinho, mas foram mantidos apenas dois pré-molares em cada meia mandíbula. Uma evidência importante da convergência com outros polinizadores da família Phyllostomidae, pois em Anoura (um morcego do clado de Vampyrum) o alongamento do focinho produziu pré-molares adicionais nas maxilas, uma neoformação comparável ao aumento de dentes dos golfinhos, de Priodontes maximus ou do crocodilo Gavialis gangeticus. "Assim como a pedomorfose pode reduzir o número de segmentos embrionários, reduzindo comprimentos. Esses segmentos podem ser aumentados pelo aumento do "timing" de crescimento na fase embrionária"
Evolução de Phyllostomus-Carollia-Sturnira e Brachyphylla

Observe atentamente, Sturnira tem incisivos, caninos e premolares como Phyllostomus e Carollia. Enquanto que em Brachyphylla + (Uroderma + Artibeus + etc) houve redução dos pré-molares e dentes da frente.
Duas evoluções independentes.....mas setorizadas meio a meio ! Sugerindo um pacote gênico !
Carollia subrufa é a mais conservadora das espécies desse gênero. O que nos limita à América Central como possível centro de origem do grupo, pois espécies se originam em um ambiente numa coevolução com esse ambiente, e estão no lugar ao qual se adaptaram. Carollia subrufa se originou ali, e na comparação com as outras espécies se mostrou a mais conservadora. Isso é muito claro ! Seguindo esse mesmo raciocínio Phyllostomus se originou na Região Amazônica ou no mínimo incluía essa região.
Mostro aqui também os centros de origem dos gêneros Sturnira e Brachyphylla ( e do grupo que reune Erophylla e Phyllonycteris com este).
Percebam que existem DUAS POLARIDADES CRUZADAS ! Phyllostomus-Carollia (com diferença principalmente de tamanho) e Sturnira-Brachyphylla (Com significativa diferença ECOLÓGICA ! Uma distribuição corta a outra bem no meio...ou seja reflete o meio... Sturnira tem tamanho intermediário entre Carollia e Phyllostomus.
"Para se determinar o lugar de origem de um grupo só existe uma maneira !
Temos que fazer uma filogenia !
Uma vez determinado dentro desse grupo qual de seus membros é o mais basal, a área de origem será a distribuição geográfica desse elemento basal, mesmo que esse elemento seja um fóssil ! O lugar onde esse fóssil foi achado, será o lugar de origem ! Obviamente se outro organismo, não conhecido na primeira análise for considerado como basal, tudo muda ! Não existe outra possibilidade !"
Segundo a Teoria dos Fractais Alométricos que venho desenvolvendo:
A EVOLUÇÃO trabalha nas pontas "dos braços" do FRACTAL EMBRIONÁRIO ! Eu vi isso nos morcegos, vocês não ?
Orelhas ...focinho....extremidades (asas e pernas ) e Cauda ! Lembram ? "A regra do gato siamês" ?
Pensem também nos ratos fossoriais....olhos e orelhas reduzidos....cauda reduzida....e assim por diante.....
é aí nessas pontas do "fractal embrionário ancestral" que acontecem as mutações e as variações genéticas..
Lá ENTRE os dentes anteriores e posteriores (bem entre os molares e os pré-molares) existe um "gerador de dentes".... produz dentes desse centro para a parte posterior e para a parte anterior...
molares e pré-molares simultaneamente.
Em termos evolutivos, o instante de parada (O CORTE PEDOMÓRFICO) vai determina a "quantidade de dentes ou seu estágio de desenvolvimento.....final".
Observe atentamente, Sturnira tem incisivos, caninos e premolares como Phyllostomus e Carollia. Enquanto que em Brachyphylla + (Uroderma + Artibeus + etc) houve redução dos pré-molares e dentes da frente.
Duas evoluções independentes.....mas setorizadas meio a meio ! Sugerindo um pacote gênico !
Vocês conseguem entender isso ?
Aparentemente existe um "fractal ancestral" totalmente compartilhado por todos os membros da espécie e MAIS as pontas que representariam a variação individual...ou seja a Variação periférica !
Nessas pontas do "fractal ancestral", ficariam as Variações Periféricas....por isso temos sempre um certo número de "defeitos congênitos" como dedos fundidos...etc, acontecendo aí.
Nessas extremidades ocorrem as mutações e os cortes pedomórficos !
Esse é o meu modelo para se pensar a evolução...tudo pode ser visto na evolução de Monodelphis e nos morcegos...
e muitas coisas virão para melhorar esse entendimento e para ajudar na compreensão dos grupos que apresentarei a seguir !
Descobri que as "variações periféricas" tem polaridades geográficas....
Por exemplo, a distribuição geográfica de tamanho do corpo no "ancestral de Phyllostomus-Carollia", provavelmente ia da América do Sul (grande) para a América Central (pequeno),
apresentando "um gradiente de tamanho" entre as extremidades geográficas. A fragmentação dessa distribuição deu 2 espécies de tamanhos diferentes e ecologias diferentes.
Ou seja, Carollia e Phyllostomus!
Veja e pense nas duas espécies de tatus que saíram de Cabassous chacoensis ...C.truncatus e C. retusus.....ocorrem justamente nas extremidades da distribuição geográfica de Cabassous chacoensis.
Adaptações costumam apresentar gradientes, pois esses gradientes são determinados pelo grau de umidade.... que é proporcional ao grau de energia disponível nos ambientes !
O tamanho do corpo, se distribui da máxima umidade ambiental (Tamanho grande) para a mínima umidade (tamanho pequeno) !
No caso de C. retusus e C. truncatus a quebra entre essas duas espécies, se deu provavelmente nas encostas das montanhas, onde outrora teria existido uma população intermediária, e que deveria ser contínua "no início" com Cabassous chacoensis!
English text :
Carollia subrufa is the more conservative of the species of this genus. What limits us to Central America as a possible center of origin of the group, because species originated in an environment in coevolution with that environment, and are in the place to which they adapted.
Carollia subrufa originated there, and in comparison with the other species showed more conservative. This is very clear!
Following this same reasoning Phyllostomus originated in the Amazon region or at least included this region.
Show here also the centers of origin of genres Sturnira and Brachyphylla (and the group gathers Erophylla and Phyllonycteris with this).
Notice that there are TWO POLARITIES CROSSED! Phyllostomus-Carollia (with differences mainly in size) and
Sturnira-Brachyphylla (With significant ECOLOGICAL difference). One distribution cuts the other right in the middle ... ie reflects the meddle of that distribution ...Sturnira has intermediate size between Carollia and Phyllostomus.
"To determine the place of origin of a group, there is a way only!
We must do a phylogeny!
Once determined within that group which of its members is the most basal.....
original area is the geographical distribution of this basal element, even if that element is a fossil! The place where this fossil was found, it is the place of origin! Obviously if another organism, not known in the first analysis is considered basal, everything changes! Does not exist another possibility! "
According to the Theory of Allometric Fractals I have been developing:
EVOLUTION works at the tips "of arms of the EMBRYONIC FRACTAL ! That I saw in the bats, do not you?
Ears ... nose .... extremities (legs and wings) and tail! Remember? "The rule of the Siamese cat"?
Consider also in mice Fossorial....eyes and ears reduced ......reduced tail and so on .....
Here it is in these tips that mutations and variations occur ..
The figure above (in the LINK) shows an interesting thing:
There BETWEEN the anterior and posterior teeth (right between the molars and premolars) there is a "generator of teeth" .... producing teeth from the center to the rear and to the front ...premolars and molars simultaneously.
In evolutionary terms, the instant stop (PAEDOMORPHIC CUT ) will determine the "amount of teeth or their stage of FINAL development."
Look closely, Sturnira have incisors, canines and premolars as Phyllostomus and Carollia. While in Brachyphylla + (Uroderma Artibeus + + etc.) there was a reduction of the premolars and front teeth.
Two independent developments ..... but sectored half and half! Suggesting a genetic pack!
Do you understand that?
Apparently there is a "ancestor fractal" fully shared by all members of the species, and the tips that represent individual variation ... ie the peripheric variation!
In these tips of the "ancestor fractal" , would Peripheric Variations .... so we always have a number of "defects" as fused fingers ... etc, happening there.
In these tips mutations occur and paedomorphic cuts !
This is my model for thinking about evolution ... everything can be seen in the evolution of Monodelphis and bats ...
and many things will come to improve this understanding and to help in understanding of the groups that presented below!
I found that "peripheric variations " has geographical polarities ....
For example, the geographic distribution of body size in the "ancestral Phyllostomus-Carollia" probably was distributed from South America (large) to Central America (small) presenting "a size gradient" between the geographical extremities. The fragmentation of this distribution originated two species of different sizes and different ecologies.
I.e., Carollia and Phyllostomus!
See and think about the two species of armadillos who detached from Cabassous chacoensis ... C.truncatus and C. retusus ..... occur axactly at the ends of the geographical distribution of Cabassous chacoensis.
Adaptations usually present gradients, since these gradients are determined by the degree of moisture .... which is proportional to the energy available in the environment!
The size of the body, distributes the maximum ambient humidity (Large Size) for the minimum humidity (small size)!
In the case of C. retusus and C. truncatus the break between these two species, probably occurred on the slopes of the mountains, where once there would have been a intermediate population, and should be continued "at the beginning" with Cabassous chacoensis!
Carollia Phylogeny (some relations) and sisters

Só para adiantar alguma coisa. Um gênero a menos nos Phyllostomidae, Rhinophylla pumilio = Carollia pumilio.
Evolução.......Sim ! Grau de evolução.......Não !
Como não poderia deixar de ser.....Na molecular......que é "gradista" como a cabeça da maioria dos BIÓLOGOS.....
"Se um organismo se modificou muito e atingiu um "NOVO GRAU" separa-se, cria-se um grupo parafilético,....e sai "LOGO DE CARA" na PHYLOGENYA"
Em breve "ALL The History" !
Vejam que comparo claramente Carollia subrufa com Rhinophylla pumilio....e logo abaixo comparo Sturnira magna com S.bidens. Em ambos os casos ressaltando uma polarização por redução de tamanho.
Prestem atenção que as semelhanças existentes entre os crânios de Rhinophylla pumilio e Sturnira bidens são típicas de "rotas" pedomórficas já vistas em outros morcegos. Vale lembrar que os dentes simplificados de Cebuella e Callithrix já foram considerados como primitivos...só uma demorada discussão abriu as portas para um nova visão. Os dentes de Rhinophylla são semelhantes (em sua simplificação) aos de alguns filostomídeos polinizadores. Mas ficaria bem difícil de considerar isso como sinapomorfias.
Ao meu ver Rhinolphylla se encaixa perfeitamente numa sequência pedomórfica dentro de Carollia e em nenhum outro lugar.
Como aconteceu em Cebuella, os dentes de Rhinophylla são resultado de simplificação....seguindo uma rota de simplificação que já se vê na base de Carollia, que por sua vez já é bem simplificado. O padrão plesio está em Phyllostomus (com bacias hipoconais evidentes). Carollia já se simplificou bastante devido a redução de tamanho, deve ter partido certamente de um modelo com amplas bacias hipoconais.
Comparation - Phyllostomus-Carollia-Rhinophylla

The progressive reduction of the molars shows a series of transformations.
The teeth are out of scale, just to compare series of transformations.
Rhinophylla is Carollia.
COMPARATION -Phyllostomus-Carollia-Rhinophylla

The polarization of some characters : All agree with a paedomorphic patterns. Mandibles in the same scale.
Cheek teeth out of scale (only comparative in size).Carollia perspicillata shows less transformation than Carollia castanea and Rhinophylla in the shape of the lower cheek teeth, when compared with Phyllostomus .
Phyllostomus is a excellent OUT-GROUP, it has the ancestral pattern of teeth.

The molar teeth reflect the type of feeding and therefore part of the ecology of these groups.
The comparison with related groups, will allow us to better understand the evolution of Stenodermatinae.
I propose with this comparison, show that Brachyphylla, Erophylla and Phyllonycteris ( subfamily : Brachyphyllinae ), are directly connected to the base of Stenodermatinae. So I'm including these genres in its natural place alongside their sisters groups.
Brachyphylla and these related genera form a monophyletic group, which apparently evolved in the West Indies. Brachypylla is more conservative than the other genres that evolved to a diet based on pollen.
Six genera are compared, Phyllostomus among them is undoubtedly the most conservative.
Sturnira is in the base of all other genres of Stenodermatinae, it still has large incisors (at least in the basal species) and developed canines, slightly modified, similar to Carollia and Phyllostomus.
Brachyphylla, Artibeus and Uroderma have incisors and canines reduced in size.
Sturnira suffered reduction and simplification of the molars, but it is possible to detect this reduction came from a model similar to other Stenodermatinae, showing an enlargement with tendency of reduction of the last molar, which is only slightly reduced in Brachyphylla.
Brachyphylla and Erophylla have very reduced P3 (first upper premolars) and second incisors, a synapomorphy that unified these genera.
Brachyphylla, Erophylla and Phyllonycteris are not included within Stenodermatinae, but this is undoubtedly its natural place.
Brachyphylla Gray was placed by Gervais (1855-1856) in his Tribe Stenodermina, apud Taboada & Pine, 1969.
* I put the left tooth of Erophylla (inverted) side by side....to show variation in the same individual....and because the right tooth is broken ! hehehe
The FRACTAL...moves from the center to the periphery....branching...branching...and branching...stretching by LOOPINGs.....shortening by CUTs.....in the TIPS....the manifestations of non LETHAL variations...survives...... allowing the existence of the EVOLUTION
Pay attention..the evolution in upper.....IS GREATer...than in lower !
HÃ ....hã...hã ???? Because.....There is independent evolution in every branch ! But the pedomorphic CUT occurs in whole sistem...stopping the whole PROCESS !
Sturnira Evolution 1

Esta prancha apresenta os grupos externos (outgroups) utilizados para a polarização de caracteres dentro do gênero Sturnira.
Na base temos Phyllostomus e Carollia, que representam dois grupos basais conservadores, sendo Phyllostomus o mais conservador deles e Carollia uma espécie de miniaturização pedomórfica do mesmo padrão apresentado por Phyllostomus. Ao meu ver, esse padrão representa uma primeira irradiação desse grupo como um todo. Parece que houve no início desse ramo evolutivo, uma grande variação no tamanho do corpo, com uma separação ecológica alimentar baseada em itens semelhantes, mas de tamanhos diferentes.
Os Sternodermatinae representam uma nova abordagem ecológica. Com dentes mais simplificados se compararmos a Phyllostomus, que é nosso padrão basal mais conservador. Para compreender melhor a evolução que aconteceu dentro de Sturnira, temos que levar em conta os principais rumos evolutivos dos Stenodermatinae.
1 - Vampyriscus (Vampyressa) possui um focinho curto e estreitado na ponta, com redução dos caninos e incisivos, além da perda do terceiro molar.
2 - Artibeus-Ametrida possuem focinhos curtos e largos, com encurtamento maior em Ametrida. Os dentes são menos simplificados que em Vampyriscus, com hipocones evidentes nos molares, mas com redução de tamanho ou perda do terceiro molar.
3 - Brachyphylla representa um outro rumo evolutivo, bem basal com crânio mais longo e dentes pouco modificados se compararmos aos demais Stenodermatinae, mas com significativa modificação nos incisivos e em P2.
Diante desse quadro geral, podemos tranquilamente, polarizar os caracteres apresentados pelas espécies de Sturnira.
Com base em Phyllostomus e Brachyphylla podemos dizer que o crânio de Sturnia teve um encurtamento no focinho, que foi além daquele já apresentado pelo ancestral dos Stenodermatinae, no entanto manteve os caninos e incisivos pouco modificados.
As espécies Sturnira magna e S. tildae se destacam por apresentarem os incisivos e caninos mais conservadores de todos os Stenodermatinae.
O pré-molar 3 superior (o primeiro dos dois pré-molares) é conservador em Sturnira, mais que nos outros gêneros, assemelhando-se a Phyllostomus.
P4 (o segundo) está simplificado em Sturnira se comparado a Brachyphylla, que mantém mais a aparência de Phyllostomus e portanto conservador.
Os molares são simplificados em Sturnira se comparamos a Brachyphylla ou mesmo a Artibeus ou Uroderma. Sturnira tildae ainda apresenta uma pequena expansão na região dos hipocones.
Com relação ao tamanho do crânio, Brachyphylla é o mais conservador de todos os Stenodermatinae, seguido por Sturnira. A crista sagital de Brachyphylla é intermedária entre Sturnira e Phyllostomus.
"Essas comparações deixam bem claro que Sturnira magna e S. tildae são mais conservadoras e as espécies menores as mais derivadas no conjunto."

Nesta prancha apresento várias espécies de Sturnira, agrupadas com base nas características cranianas. Os grupos irmãos mais imediatos e conservadores (Phyllostomus e Brachyphylla) nos ajudam nessa visualização.
Observem as setas vermelhas que indicam algumas sinapomorfias e os ângulos formados pelo alinhamento incisivos-canino (ângulo formado em relação ao eixo do crânio e as cristas do canino e do inscisivo 2, como se vê na figura)
Com base nessas espécies é possível construir um esboço da evolução biogeográfica do gênero.
De acordo com o raciocínio que tenho apresentado ao longo de minhas postagens, a espécie mais conservadora está no centro de origem.
Portanto Sturnira magna, que está distribuida pela vertente oriental dos Andes, desde a Bolivia até a Colombia, se conservou por estar em seu lugar de origem.
Seguindo o rumo proposto pela sequência da filogenia, temos uma expansão a partir dessa área, originando Sturnira tildae e S. angeli com distribuições complementares (vicariantes).
Sturnira mordax deve ter se originado na América Central junto com S. angeli, pois derivaram conjuntamente (talvez até de um ancestral comum, mas S. angeli é preferncial, devido a sua distribuição).
Pelo fato desse par estar mais relacionado com as demais espécies, podemos pensar numa expansão de volta para a América do Sul entrando em simpatria com as espécies mais antigas.
Sturnira erythromos e S. lilium formam outro par com uma complementaridade parcial entre si, entrando em simpatria com S. magna e S. tildae de "maior tamanho" talvez ocupando outro nicho sem competição.
Sturnira ludovici e suas complementares S. bogotensis e S. oporaphilum, representam uma irradiação andina com expansão de S. ludovici para a América Central. Essa quebra em 3 unidades é muito semelhante as quebras de Mazama.
Sturnira bidens e S. nana parecem representar uma irradiação terminal ocupando as maiores altitudes dos Andes setentrionais onde prevalece a umidade.
Infelizmente essa filogenia parcial das espécies não permite maiores comentários.
Sturnira Biogeography

O gênero Sturnira se originou a meio caminho de uma distribuição contínua (Carollia-Phyllostimus). Possui um tamanho intermediário e mescla características intermediárias, como uma folha nasal reduzida e um focinho curto.
Talvez uma adaptação às montanhas.
Inicialmente se deslocou para o sul ao longo dos Andes, indo até os limites tropicais (padrão que se observa em inúmeras espécies). Provavelmente representou a quebra da distribuição inicial Carollia-Phyllostomus.
Num segundo tempo entra em simpatria tanto com Carollia e como com Phyllostomus. O que acabou resultando em duas outras linhagens. Essas linhagens refletem o padrão inicial, tamanhos diferentes proporcionais aos ambientes. A linhagem menor na América Central (como Carollia) e a maior Amazônica (como Phyllostomus).
A linhagem menor ( S. mordax e S. angeli) se expande para o ambiente "mais rico", entrando em simpatria com as outras espécies.
Mais duas linhagens se formam :
1 - Sturnira erythromos e S. lilium
2 - Sturnira ludovici e suas espécies irmãs S. bogotensis e S. oporaphilum (evolução paralela à do gênero Mazama).
Essa análise necessitaria de mais informações, sobre as espécies não relacionadas.
The genus Sturnira originated halfway from a continuous distribution (Carollia-Phyllostimus). It features a intermediate size and merges intermediate characteristics, as a leaf nose reduced, and a short muzzle. Perhaps an adaptation to the mountains.
Initially moved south along the Andes, going to the limits tropical (pattern that is observed in many species).
Probably represented "the breaking" of the initial distribution Carollia-Phyllostomus.
In a second time enters into sympatric with both Carollia and as with Phyllostomus. What eventually resulted in two other lineages. These lines reflect the standard initial sizes proportional to the environments. The minor lineage in Central America (as Carollia) and greater Amazon (as Phyllostomus). The minor lineage (S. mordax and S. Angeli) expands to the environment "richer", entering sympatric with other species. Two more lines are formed:
1 - Sturnira erythromos and S. lilium
2 - Sturnira Ludovici and its sisters species S. bogotensis and S. oporaphilum (parallel to the evolution of the genus Mazama).
This analysis would need more information about unrelated species.

A principal modificação, observada nesse grupo de morcegos filostomídeos, é o alargamento do palato, culminando nos gêneros mais modificados, com um grande encurtamento.
Podemos observar dois grupos principais. Um deles é mais conservador, com morcegos pouco diferenciados, a partir de um tipo Artibeus. O outro é formados por morcegos muito interessantes, apresentam olhos saltados e boca larga. Nesse grupo estão dois dos morcegos neotropicais mais extravagantes, Centurio e Sphaeronycteris. Ambos possuem o focinho e o palato muito encurtados. Tudo indica que o encurtamento do palato aconteceu duas vezes, pois Ametrida e Sphaeronycteris possuem crânios muito peculiares, com uma caixa craniana globosa e o focinho extremamente reduzido. Pygoderma e Centurio possuem mandíbulas bem encurtadas, com o processo angular bem reduzido, que são as principais características cranianas sinapomórficas.
No primeiro grupo temos na base Phyllops, pouco modificado mas apresentando já um encurtamento da mandíbula que é também a marca do grupo. Ariteus tem um avanço no encurtamento e Stenoderma um encurtamento bem peculiar no palato, tanto na região anterior como na posterior.
A figura não mostra o gênero Ardops, que está provavelmente mais associado a Ariteus pela redução mais acentuada da mandíbula. Temos uma heterobatmia na base de Stenoderma e Ariteus-Ardops. Redução extrema do palato em Stenoderma e redução extrema da mandíbula nos outros.
A escolha de Artibeus como grupo irmão, é bastante óbvia, uma vez que dentre os filostomídeos de palato curto e largo, esse gênero é o mais plesiomórfico, representando um tipo intermediário, entre Stenodermatina e os demais. Certamente deve ser tratado também como parte desse grupo monofilético, já que é o gênero diretamente ligado aos outros 7 gêneros.
Portanto proponho que Artibeus seja tratado como Sternodermatina.
A prancha é tão clara e simples, que fica fácil em grande aumento, visualizar outras sinpomorfias, que não apontei, mas que são gritantes. Quando da publicação final todas serão mencinadas devidamente.
NOCTILIONOIDEA (Skull and rhinarium)

Na figura acima temos uma comparação de crânios e de rinários.
Na primeira série acima vemos crânios de Vampyrum, Megaderma e Myotis.
Os dois primeiros são morcegos tropicais de porte médio com folhas nasais ao redor do rinário. São rinários bem mais caraterísticos e interessantes [derivados]. Nesta figura mostro as folhas nasais de Lophostoma e Megaderma.
Myotis possui um rinário mais conservador, mas com narinas praticamente em contato com a boca. Veja em baixo a direita um rinário de Pteropodidae que poderá servir de modelo de conservadorismo [primitividade] dentro da evolução dos Chiroptera. Plecotus é mais conservador do que Myotis, mas apresenta uma expansão na parte posterior das narinas em contato com as glândulas que contornam o rinário. No “outgroup” temos também Myzopoda que apresenta um rinário semelhante a Myotis, porém mais simplificado [as narinas são quase redondas] como é comum dentro dos Nataloidea.
Na parte inferior da figura temos 3 crânios de NOCTILIONOIDEA mostrando as modificações progressivas que existem entre eles. As caraterísticas marcantes e também sinapomorfias, desse grupo formado pelas famílias Mystacinidae, Mormoopidae e Noctilionidae, são as modificações que marcam todo o focinho desde a ponta que é achatada e relativamente alta, até a curvatura pronunciada na região interorbital.
Essa modificação do focinho se reflete no rinário, que apresenta narinas relativamente próximas de forma quase tubular. Aparentemente Noctilio ainda retém uma condição onde o tubo da narina não se fechou totalmente como acontece em Mystacina. Pteronotus têm as narinas tubulares incorporadas ao rinário que é bem mais modificado, apresentando uma projeção semelhante à uma folha nasal, posteriormente. Em Mormoops o rinário atinge um grau maior de modificação que Pteronotus.
Superfamilia Noctilionoidea ( Noctilionidae, Mormoopidae e Mystacimnidae )
Noctilionidae e Mormoopidae são da America tropical e Mystacinidae da Nova Zelãndia.
Estou dividindo os Chiroptera em 6 superfamílias:
PTEROPODOIDEA
PHYLLOSTOMOIDEA
RHINOLOPHOIDEA
NOCTILIONOIDEA
NATALOIDEA
VESPERTILIONOIDEA
Essas superfamílias se distribuem em 3 GRANDES GRUPOS :
subordem : Megachroptera ( Pteropodoidea )
subordem : Microchiroptera:
- Infraordem : Noctiliformes ( Phyllostomoidea+Rhinolophoidea+Noctilionoidea )
- Infraordem : Vespertiliformes ( Vespertilionoidea+Nataloidea )
Origem e dispersão dos CHIROPTERA


Primeiramente vamos tratar da questão mais importante, referente ao mapa, que aqui apresento. Como este mapa foi construído ?
Obviamente este mapa é hipotético.
Sua construção se baseia em certos princípios, que venho trabalhando, e estão presentes em muitas das minhas postagens.
1 Conservação de caracteres plesiomórficos,
2 Distribuição biogeográfica gradativa de caracteres,
3 Distribuição geográfica associada a evolução desses caracteres citados.
Os caracteres plesiomórficos estão associados as áreas de origem
e os derivados [apomórficos] às áreas de dispersão posteriores.
Poderíamos dizer que a evolução dos caracteres reflete a dispersão de um determinado grupo. Pois o ambiente modela os seres vivos.
Ambientes conservadores preservam antigos caracteres e ambientes modificados transformam esses caracteres.
1 - Dermoptera é o grupo-irmão de Chiroptera, as evidências são muito boas, embriológicas e anatômicas, principalmente. Esses animais têm uma distribuição geográfica restrita, vivem só nas florestas do sudeste asiático. Possuem uma alimentação à base de vegetais e seus dentes não se parecem com os dentes mais primitivos, encontrados em morcegos, insetívoros e marsupiais. Seu crânio se modificou em relação ao provável padrão ancestral dos mamíferos, e essas modificações foram principalmente nos dentes, indicando que a alimentação desses animais é um tipo derivado e especializado. A locomoção por planeio deve ter dado origem ao voo dos morcegos. Os morcegos passam por uma fase embrionária, que se assemelha bastante ao padrão de membranas, encontrado em Dermoptera.
2 – Pteropodídeos são morcegos mais conservadores com relação ao tamanho e caracteres gerais, suas modificações mais evidentes, assim como vemos em Dermoptera, são os dentes. Podemos dizer que também como aqueles, suas mudanças foram com relação ao tipo de alimentação.
3 – Os microquirópteros representam a grande maioria dos morcegos e sua distribuição é mundial, ocupando todo o planeta, com exceção da Antártida. Seus dentes refletem a alimentação primitiva dos morcegos, INSETIVORIA.
Por que Tarsius [que é o grupo-irmão dos primatas superiores] está no SUDESTE ASIÁTICO ? Assim como Tupaídeos....
Por que também os quirópteros mais conservadores [Pteropododae] e Dermoptera também estão ali ? e Echinosorex com suas características conservadoras ?
Os pteropodídeos africanos são derivados dos asiáticos e formam um grupo visivelmente homogêneo, "com suas manchas brancas na base das orelhas".
Ali se conservaram as antigas faunas florestais do planeta.
Os morcegos se expandiram numa fase úmida para as outras regiões do planeta... mas sofreram uma grande extinção advinda de uma secura generalizada.
Essa secura explica a redução do tamanho dos Microchiroptera, e as transformações drásticas que sofreram as floras e faunas da África [veja RAVEN, P.H. & AXELROD, D.I. 1974. Angiosperm Biogeography and Past Continental Movements.]
As florestas do sudeste asiático representam uma região onde sempre houve uma grande estabilidade climática.
Ali nessa região..... mesmo nas fases climáticas mais secas, nas quais as outras florestas sofreram grandes transformações [África e Américas], nunca houve uma grande transformação !
Pelo contrário aquela fauna é marcada por grandes fases de alagamento que resultou em um tipo de evolução relacionada aos "planadores"........cobras planadoras...rãs planadoras...esquilos, geckonídeos, as adaptações do gênero Draco (dragão-voador), origem dos quirópteros....
No final do CRETÁCEO não existiam os mesmos oceanos que conhecemos hoje, portanto não existiam os mesmos fatores climáticos que existem hoje !
O único OCEANO [atestado pela antiguidade dos sedimentos marinhos] era o Pacífico norte....ÚNICO!
Portanto a única floresta com capacidade de resistir as fases secas era BEM ALI, no sudeste asiático !
O giro da TERRA leva sempre chuvas onde as correntes marítimas batem de frente. Mesmo nas épocas mais secas, Madagáscar, Nova Zelândia e Nova Guiné e Austrália sempre conservaram uma costa oriental úmida. Isso explica por que nessas regiões existem uma séries de formas conservadoras, tanto no reino vegetal como no animal.
ESTABILIDADE é a palavra !
Se pensarmos no PLANETA no final do Cretáceo, teremos um quadro exatamente desse jeito...chuvas sempre presentes no sudeste asiático, provenientes das correntes marítimas do ÚNICO OCEANO da época.
Essas chuvas mantiveram estas florestas quase intactas e sua fauna foi preservada.
As florestas Africanas herdaram a sua fauna da PRIMITIVA FLORESTA ASIÁTICA.....
Certamente houve uma dispersão da fauna asiática em direção à África, em uma fase úmida do planeta... Isso inclui Primates e Rodentia.
Mas quando ocorreu a fase seca... só mesmo as chuvas provocadas pela "Corrente Norte PACÍFICA" puderam manter as faunas pouco modificadas, no sudeste asiático.
Os pteropodídeos "já modificados" e "com dentes derivados" que habitam a África, só colonizaram essas matas, em uma próxima fase úmida.
Ou seja, depois do aparecimento da Grande Floresta Asiática, houve a primeira expansão.
Numa segunda expansão, dispersaram-se os pteropodídeos, que muito provavelmente nem existiam na primeira expansão,
Os primeiros morcegos que se dispersaram para a África, caracterizavam-se necessariamente, por possuírem dentes como nos Phyllostomidae.
A redução progressiva do corpo, observada nos morcegos, permitiu a sobrevivência na pobreza ambiental que se instalou !
Os Megachiroptera devem ter se originado no sudeste asiático, enquanto os Microchiroptera mais provavlemente se originaram na África, dispersando-se para a América do sul. Os Phyllostomidae são os Microchiroptera mais conservadores [particularmente Vampyrum e Phyllostomus].
Let us first deal with the most important question, concerning to the map, which I present here. How this map was built?
Obviously this map is hypothetical.
Its construction is based on certain principles, which I have worked, and are present in many of my posts.
1 Conservation of plesiomorphic characters.
2 Biogeographical distribution of progressive characters.
3 Geographical distribution associated with the evolution of these characters mentioned.
The plesiomorphic characters are associated to origin areas
and derivatives [apomorphic] to further dispersal areas.
We could say that the evolution of the characters reflects the dispersion of a particular group, since the environment modeling the living beings.
Conservatives environments preserve old characters and modified environments transform these characters.
1 - Dermoptera is the sister group of Chiroptera, the evidences are very good, embryological and anatomical mainly. These animals have a narrow geographic range, living only in the forests of Southeast Asia. They have a feeding based on vegetables and their teeth do not look like those most primitive teeth, found in bats, insectivores and marsupials.
Its skull was modified in relation to the probable ancestral pattern of mammals, and such modifications were mainly on the teeth, indicating that the feed of these animals
is a derived type and specialized. The locomotion by gliding should have led to the flight of bats. Bats pass through a embryonic stage, which is quite similar to the pattern of membranes, found in Dermoptera.
2 - Pteropodídeos bats are more conservative with respect to size and general characteristics, their most obvious changes, as we see in Dermoptera are the teeth. We can also say that like those, its changes were related to the type of food.
3 - The microchiropteran represent the vast majority of bats and their distribution is worldwide, occupying the entire planet, except Antarctica. Their teeth reflect the primitive feeding of the bats, insectivorous.
Why Tarsius [which is the sister group of higher primates] is in SOUTHEAST ASIA? As Tupaídeos ....
Why also the more conservative bats [Pteropododae] and Dermoptera
Also are there? and Echinosorex with its conservative characteristics?
The African pteropodídeos are derived from Asian and form a group clearly homogeneous, "with its white spots at the base of the ears."
There preserved up the ancient forest faunas on the planet.
Bats has expanded in a wet phase to the other regions of the planet ... but suffered a large extinction due to an generalized dryness.
Such dryness explains the reducing size of microbats, and the dramatic transformations that have undergone the floras and faunas of Africa [see RAVEN, PH & Axelrod, DI 1974. Angiosperm Biogeography and Continental Movements Past.]
The forests of Southeast Asia represent a region where there has always been a very stable climate.
Here, in this region ..... even at the driest climate in which other forests have suffered great transformations [Americas and Africa], there has never been a great transformation!
On the contrary, that fauna is marked by phases of flooding which resulted in a kind of evolution related to "gliders" ........ soaring snakes, soaring frogs and. squirrels, geckonids, the glinding adaptations of the genus Draco (flying lizards), and the origin of the bats ...
At the end of CRETACEOUS there were no oceans as we know them today, so there were not the same climate factors that exist today!
The only OCEAN [attested by ancient marine sediments] was the North Pacific .... UNIQUE!
So the only forest with capacity to resist drought stages was JUST THERE, Southeast Asia!
The spin of EARTH brings rains always where the sea currents clash. Even in the driest seasons, Madagascar, New Zealand and New Guinea and Australia has always retained a moist east coast. This explains why these regions there are a series of conservative forms, both in the plant kingdom and the animal.
Stability is the word!
If we think about PLANET at the end of the Cretaceous, we have a picture exactly like this ... rain always present in Southeast Asia, ocean currents from the UNIQUE OCEAN of the time.
These rains kept these almost untouched forests and their fauna was preserved.
African forests have inherited their fauna of the EARLY ASIAN FOREST .....
Certainly there was a dispersal of Asian fauna towards Africa, in a wet phase of the planet ... This includes Primates and Rodentia.
But when the dry phase took place ... only the rainfall caused by the "North Stream Current" could kept faunas slightly modified, in Southeast Asia.
The pteropodídeos "already modified" "with teeth derivatives" that inhabit Africa, only colonized these forests in a forthcoming wet phase.
That is, after the emergence of the Great Forest Asian, there was the first expansion.
A second expansion, the dispersed pteropodídeos, that probably did not even exist in the first expansion.
The first bats dispersed to Africa, were characterized necessarily by having teeth like the Phyllostomidae.
The progressive reduction of body observed in bats, allowed survival in poverty environment which was installed!
The Megachiroptera must have originated in Southeast Asia, while Microchiroptera
most likely originated in Africa, dispersing to South America. The Phyllostomidae Microchiroptera are more conservative [particularly Vampyrum
and Phyllostomus].
Dispersion of the Plecotidae, Antrozoidae and Histiotidae

We SEE HERE the first major dispersal of Chiroptera.
The distribution of Plecotidae is very suggestive, almost restricted to temperate regions [or mountainous America] unlike Myotidae, Vespertilionidae, Emballonuridae and Molossidae, which spread widely.
This branch of the phylogeny of Chiroptera is a group that moved early in an environment that was not previously inhabited by other bats, standing out the group that has given Myotidae.
Probably dispersed together and it would explain the wide dispersal and diversification of the genus Myotis [worldwide], because it is a very old event. I consider this distribution a STRONG evidence of which my phylogeny is on course.
The Plecotidae seem to ecological constraints since its appearance.
They prefer cooler regions or desert.
I put marks of ocean currents and dry air currents sweeping Asia to reach the Sahara.
These dry air currents breaking the distributions of the moist tropical forests during dry periods of global climate.
Note also the axis of inclination of the distributions in Asia.
See how Histiotidae came to occupy the available areas not occupied by tropical bats! They live in dry and cool range of South America and the Andes Mountains, which are thus due to the altitude!
If you look at my board Evolution of Chiroptera - skulls,
will find that Plecotidae and its sister group, experienced a significant reduction in size!
Perhaps the size increase observed later in Vespertilionidae and Emballonuridae, is related to a "BACK" to the regions richest in food! [ Saccolaimus, the greatest emballonurid gender and and Scotophilus, a big vesper, occupied the most humid areas of tropic regions of Africa-Asia].
Since food availability, allows an evolution towards an increase in size. The opposite of what happens in lack of food.
VEMOS AQUI a primeira grande dispersão dos CHIROPTERA !
A distribuição de Plecotidae é bastante sugestiva, praticamente se restringe às regiões temperadas [ou montanhosas da América] diferentemente de Myotidae, Vespertilionidae, Emballonuridae e Molossidae, que se espalharam amplamente.
Esse ramo da filogenia dos CHIROPTERA é um grupo que precocemente avançou em um ambiente, que antes não era habitado por outros morcegos, se destacando do grupo que deu Myotidae.
Provavelmente se dispersaram juntos e isso explicaria a grande dispersão e diversificação do gênero Myotis [mundial], por ser um evento muito antigo.
Considero essa distribuição uma FORTE evidência de que a minha filogenia está no rumo certo.
Os Plecotidae parecem ter restrições ecológicas desde seu surgimento.
Preferem regiões mais frias ou desérticas.
Coloquei marcas das correntes oceânicas e das correntes de ar secas, que varrem a Ásia até chegar ao Sahara. Essas correntes de ar secas quebram as distribuições das Florestas Tropicais úmidas nos períodos secos do clima global.
Observe também o eixo de inclinação das distribuições na Ásia.
Vejam como Histiotidae veio ocupar as áreas disponíveis, não ocupadas pelos morcegos tropicais ! Vivem na faixa seca e fria da América do Sul e nos Andes, que são assim devido à altitude !
Se você olhar minha prancha de Evolução de CHIROPTERA - crânios, verá que Plecotidae e seu grupo-irmão, tiveram uma redução significativa de tamanho !
Talvez o aumento de tamanho observado posteriormente em Vespertilionidae e Emballonuridae, esteja relacionado à uma "VOLTA" às regiões mais ricas em alimento !
Pois disponibilidade de alimento, permite uma evolução no sentido de uma aumento de tamanho. O oposto do que acontece na carência de alimento.
Mapa Microchiroptera tropical

Em seu quadro evolutivo os CHIROPTERA estritamente tropicais, representam uma etapa intermediária entre os Pteropodídae e os Microchiroptera mais derivados, como os VESPERTILIONIDAE, MOLOSSIDAE, RHINOPOMATIDAE, EMBALLONURIDAE e as famílias menores.
Os Phyllostomidae são exclusivos da América tropical representados em VERDE e os Rhinolophoidea [Megadermatidae, Nycteridae e Rhinolphidae] habitam o velho mundo e a Austrália.
A distribuição dos Nycteridae é disjunta assim como a dos Megadermatidae, interpreto essa quebra na distribuição como sendo produto de uma grande transformação ambiental na região central.
As regiões centrais, que hoje são em grande parte desérticas, certamente sofreram um ressecamento após um período de umidade, que permitiu a ligação entre as florestas africanas e asiáticas.
Os Rhinolophidae que considero o grupo-irmão [mais derivado] dos Nycteridae, devem ter se originado nessas áreas intermediárias, uma vez que possuem uma distribuição avançada, que ultrapassa a faixa tropical propriamente dita.
Os demais Microchiroptera citados acima como mais derivados, evoluíram no sentido de ocupar áreas mais setentrionais, portanto mais frias, distanciando-se assim das áreas de origem dos CHIROPTERA mais basais.
No entanto assim como aconteceu com os Rhinolophidae, a expansão dos grupos mais derivados, não se restringiu as áreas setentrionais, mas acabaram ocupando também as regiões tropicais entrando em simpatria com os chiroptera mais basais e estritamente tropicais.
The Ecological Evolution of Rhinolophoidea is quite clear. This group is restricted to the old world. Unlike other Microchiroptera, they never reached the latitudes near the north pole, as others who managed to disperse to America. The Megadermatidae and Nycteridae being limited to warmer regions. The Rhinolophidae (sensu lato) originated in the region of "breaking" of tropical forests. After savannization and subsequent desertification of these regions, which separate the tropical forests, the groups of forest was separated and Rhinolophidae have adapted to these intermediate regions. The absence of forests has reduced the number of shelters and Rhinolophidea along with Emballonuridae and Rhinopomatidae (in a PARALLEL EVOLUTION) used the caves as shelter. This led to an evolution in a convergent manner (echolocation and consequent anatomical changes), as these animals are often found together in the caves living in the same environments. This fact
was noted by the earliest researchers of bats.
After this initial adaptation to the modified intermediate regions, those derivatives groups dispersed to the tropical forest, thus undertaking a new evolutionary stage !
EVOLUTION in RHINOLOPHOIDEA

The Rhinolophoidea had its evolution largely confined to warm regions of the Old World (with little penetration in the temperate zone).
They are divided into 3 subgroups ( families ) :
Megadermatidae (depicted here by Macroderma), Nycteridae (Nycteris) and Rhinolophidae (Hipposideros).
The family Megadermatidae is the most conservative with respect to the size, general characteristics and behavior. The other 2 families show significant changes as a reduction in size and other paedomorphic characteristics such as eye reduction and changes in nasal leaf.
The family Nycteridae retained big ears of the Megadermatidae, but suffered a drastic reduction in eye size and larger change in nasal leaf and nostrils, a paedomorphosis comparable to certain anomalies of our species, as the malformation known as "harelip" (associated with absence of nasal septum and separation of the incisors in a central unit).
The family Rhinolophidae suffered reduction in the size of the ears, in nasal leaf and eyes (most evident in the genus Rhinolophus). This great difference in Rhinolophus generated the split of this group in a separate family (a gradistic vision that always separates everything is different and seems to have no end) thus creating a paraphyletic family Hipposideridae. The skull of Rhinolophidae is characterized by having an inflated and higher rostrum, than the other Rhinolophoidea.
The sister GROUP of Rhinolophoidea is Phyllostomoidea, they share a specialized nasal leaf around the nostrils, which is the most characteristic mark of these two groups. The rinarium is moved up, with an upper "lip" wide. Note in the skull the nasal aperture dorsally displaced.
See the homologous structures of the nasal leaf .
The genus Vampyrum is the most conservative of all the genres of Microchiroptera.
Its size differs very little from that of the largest genera of Megachiroptera.
Its skull and brain are very little modified compared to large bats.
The nasal leaf is the main modification presented by Vampyrum besides the size reduction of the body.
"The evolution of Microchiroptera should be associated ( in its origin ) to a night life (for presenting reduction of eyes, big ears and some kind of nasal modification, either internal or external).
The Noctiliformes or have a nose leaf, or nostrils formed by tubes, and the Vespertiliformes have an internal change in the muzzle with a diastema between the incisors, also associated to big ears like those of Plecotidae".
Rhinarium - Synapomorphic Nose Leaf

Sinapomorfia indiscutível compartilhada pelos Phyllostomoidea + Rhinolophoidea.
Observe os grupos irmãos e veja como o rinário está totalmente ligado à boca.
Nesses dois grupos existe uma modificação nos lábios, tanto no superior como no inferior.
Houve um afastamento do rinário para uma região mais dorsal e surgiu uma área ampla com pequenas papilas, única entre os Chiroptera.
Comparando-se os crânios podemos perceber a modificação que existe nesses dois grupos. A abertura nasal está voltada para cima (veja o ângulo).
ISSO É HISTÓRICO na classificação dos CHIROPTERA !
Na figura acima temos:
1 - foto de Vampyrum (cabeça do animal e crânio) representando Phyllostomoidea.
2 - Megaderma lyra (cabeça do animal e crânio) representando Rhinolophoidea.
3 - Os grupo externos usados como referência, Pteropodidae, Plecotidae, Myzopodidae, e outros mamíferos relacionados aos Chiroptera, Tupaia e Galeopterus, mostrando o padrão ancestral plesiomórfico.
COMPARE COM OS RINÁRIOS DE NOCTILIONOIDEA.
Chrotopterus-Rhinolophus HomologiaS

A Teoria dos fractais alométricos e a PAEDOMORPHOSIS ! As variações periféricas e seus efeitos residuais !
Como uma grande Árvore da ViDa....um grande fractal alométrico !
Com linhas definidas ! Ramos interrompidos.....geram interrupções graduais...com efeitos residuais...
A linha do olho...afeta a linha do olho....a linha da orelha afeta a linha da orelha....os lábios fendidos..as estruturas duplicadas e triplicadas....LOOPINGs.....séries...metâmeros...ramos e mais ramos de interferência !
Abrindo e fechando LEQUES !
Deslocando e expandindo...se abrindo como flores e estróbilos !
Integrados na dinâmica planetária ! As regras da PHYSIS !

RHINOLOPHIDAE EVOLUTION

Rhinolophoidea EVOLUTION - Skulls

EVOLUTION of the nasal leaf in RHINOLOPHIDAE

Rhinolophidae Evolution 2


Hill's Revision of Hipposideros

I take this Phylogeny from Hill's publication (print screen).
My view is based on my recent publications on FLICKR, that is my official vehicle of communication with the "BAT COMMUNITY" !
Hipposideros Phylogeny (simplified)


 I. PRIMATES.
I. PRIMATES.



Thomas H. Huxley em 1872, no seu Manual de Anatomia dos Animais Vertebrados, comparou anatomicamente os grupos de Insectivora estabelecidos naquela época, entre estes os Tupayae, Galeopithecus, musaranhos e toupeiras. Huxley foi o primeiro zoólogo a afirmar que as tupaias são os Insectivora que mais se aproximam dos lêmures. Ele também deu um enfoque todo especial a Galeopithecus, aproximando-o dos morcegos, considerando suas relações com os primatas "gerais e superficiais". Por fim, ele concorda com Peters, que considerou que Galeopithecus não pertence nem aos primatas, nem aos quirópteros, mas que é um Insetivora aberrante. Em suas comparações anatômicas, Huxley também ressalta que Macroscelides apresenta o rádio e a ulna anquilosados [fundidos], sem integrar essa observação adequadamente no contexto geral. Apesar dos zoólogos europeus utilizarem uma nomenclatura comum, calcada no Systema Naturae de Linnaeus, seus trabalhos ainda eram muito individuais e isolados, pois em 1811 [61 anos antes], Illiger já havia proposto a Ordo Volitantia, que reunia Chiroptera e Dermoptera, e estes últimos já estavam isolados em uma família própria. Além disso, em 1855 [17 anos antes], como citei logo acima, Wagner já havia colocado as tupaias na famíla Scandentia dentro da Ordo Insectivora, que incluía também a família Dermoptera. No entanto, Thomas Huxley, em seu isolamento, não comentou nenhuma destas publicações. Vejam abaixo os comentários dele no seu Manual de Anatomia dos Animais Vertebrados, e observem os detalhes do tratamento que ele dá às suas comparações:
__________________________________________________________________________________________










Os importantes trabalhos de Salton & Sargis.
________________________________________________________________________________________


________________________________________________________________________________________
 TERRESTRIA (Gomes, 2015) : Superorder Hypoconodonta (Gomes, 2015) Caecoidea (Gomes, 2015)
TERRESTRIA (Gomes, 2015) : Superorder Hypoconodonta (Gomes, 2015) Caecoidea (Gomes, 2015)


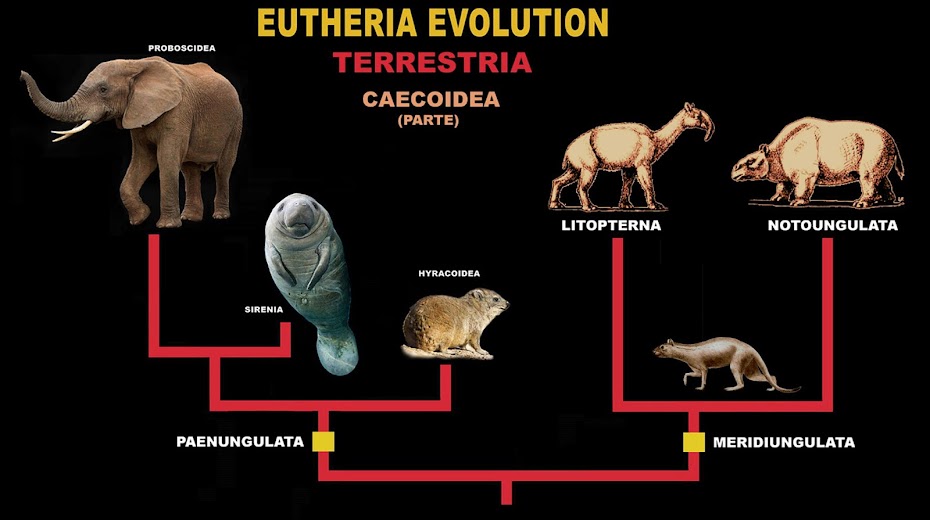
%20and%20Agnolin%20and%20Chimento%20(2011).jpg)

.jpg)
%20FOOT.jpg)
Observe os grupos irmãos e veja como o rinário está totalmente ligado à boca.
Nesses dois grupos existe uma modificação nos lábios, tanto no superior como no inferior.
Houve um afastamento do rinário para uma região mais dorsal e surgiu uma área ampla com pequenas papilas, única entre os Chiroptera.
Comparando-se os crânios podemos perceber a modificação que existe nesses dois grupos. A abertura nasal está voltada para cima (veja o ângulo).
ISSO É HISTÓRICO na classificação dos CHIROPTERA !
Na figura acima temos:
1 - foto de Vampyrum (cabeça do animal e crânio) representando Phyllostomoidea.
2 - Megaderma lyra (cabeça do animal e crânio) representando Rhinolophoidea.
3 - Os grupo externos usados como referência, Pteropodidae, Plecotidae, Myzopodidae, e outros mamíferos relacionados aos Chiroptera, Tupaia e Galeopterus, mostrando o padrão ancestral plesiomórfico.
COMPARE COM OS RINÁRIOS DE NOCTILIONOIDEA.
Chrotopterus-Rhinolophus HomologiaS

A Teoria dos fractais alométricos e a PAEDOMORPHOSIS ! As variações periféricas e seus efeitos residuais !
Como uma grande Árvore da ViDa....um grande fractal alométrico !
Com linhas definidas ! Ramos interrompidos.....geram interrupções graduais...com efeitos residuais...
A linha do olho...afeta a linha do olho....a linha da orelha afeta a linha da orelha....os lábios fendidos..as estruturas duplicadas e triplicadas....LOOPINGs.....séries...metâmeros...ramos e mais ramos de interferência !
Abrindo e fechando LEQUES !
Deslocando e expandindo...se abrindo como flores e estróbilos !
Integrados na dinâmica planetária ! As regras da PHYSIS !
Note that Rhinolophus has greatly reduced eyes ... and if you have not noticed,
exactly in line with the eyes (which have declined) we have a localized reduction in leaf nose ... right there ... see that the leaf suffered a retraction a stop right there in line with the eyes ... in the formation of the embryo, there are orders that follow one after the other ... "EMBRYONIC INDUCTION" ... one structure interferes with the formation of another structure .... in a sequence.
An order goes to formation of the upper lip and "the same order" goes to the eyes,
there in the "Eye Line" .....it affects everything that is on that particular induction! If you look at a photo, Rhinolophus also has small ears and in the "line of ears" there also exists a retraction located (in the leaf). Try to understand this!
Think of a tree growing! To be the branch tip, had to exist before the basal branch !
Observem que Rhinolophus tem os olhos muito reduzidos...e se ainda não perceberam......exatamente na linha dos olhos (que sofreram uma redução) temos uma redução localizada na folha nasal...exatamente ali... vejam que a folha sofreu uma retração uma parada bem ali na linha dos olhos...na formação do embrião, existem ordens que se sucedem uma após outra...a "INDUÇÃO EMBRIONÁRIA"...uma estrutura interfere na formação de outra estrutura....numa sucessão. Uma ordem parte para a formação do lábio superior e essa mesma ordem parte para os olhos, na linha dos olhos.....afeta tudo que está ligado nessa determinada indução ! Se observarem em uma foto, Rhinolphus também tem orelhas reduzidas e na linha da orelhas também existe uma retração localizada. Tentem entender isso !
Pensem numa árvore crescendo ! Para existir o galho da ponta, antes teve que existir o galho basal !
Chrotopterus-Rhinolophus HomologiaS 2

"O crânio dos mamíferos (e dos vertebrados de modo geral) foi formado por segmentos que se fundiram. Os nervos cranianos são as evidências desses segmentos. Durante a indução embrionária esses segmentos não são formados simultaneamente. Quando um segmento A está num estágio avançado de formação, um outro segmento B já inicia seu desenvolvimento, antes do término de A. A ponta do focinho dos Rhinolophidae revela essa ordenação. Forma-se uma estrutura semelhante a uma pirâmide, por uma questão de estabilidade do sistema. Muitas das convergências observadas na natureza são condicionadas por fatores estruturais.....(tudo uma questão de estabilidade estrutural)".
ARCANO Nelson Gomes
"Os Rhinolophidae ( através de suas folhas nasais ) nos fornecem inúmeros elementos que podem elucidar a ordem que existe na indução genética"

Inicio hoje uma série de Pranchas explicativas sobre a EVOLUÇÃO dos RHINOLOPHIDAE
Irei demonstrar nesses próximos dias que o gênero Rhinolophus está proximamente relacionado aos gêneros Triaenops e Rhinonycteris. Através de uma análise comparativa dos crânios e das folhas nasais, concluí que a estrutura conhecida como SELLA dos Rhinolphidae (sensu strictu) tem correspondências com outras estruturas das folhas nasais de Triaenops e Rhinonicteris.
Je vais vous montrer ces prochains jours, que le genre Rhinolophus est étroitement connexe aux genres Triaenops et Rhinonicteris. Travers une analyse comparative des crânes et des feuilles nasales, j'ai conclu que la structure connue comme SELLA de Rhinolphidae (sensu strictu) correspond à d'autres structures des feuilles nasales de Triaenops et Rhinonycteris.
Temos à esquerda 4 fotos de Paratriaenops furculus e à direita 2 de Rhinolophus smithersii.
Compare cada uma das partes das folhas nasais e descubra por si mesmo essas homologias, tudo foi colocado de maneira adequada para facilitar. Os pontos centrais estão indicados pelas setas, mas sempre podemos ir além !
Nesta figura tento transmitir para aqueles que podem ver, os segredos da ordenação embrionária. Essas folhas nasais são reveladoras. Revelam a ordem da indução embrionária. Através do conceito que denominei "Fractal Alométrico" tento mostrar que a ordem obrigatória dessas induções determinam convergências obrigatórias e elucida os mecanismos genéticos.
Rhinolophoidea EVOLUTION - Skulls

Skulls in the same scale. Comparative size.
Estão faltando alguns gêneros nessa filogenia. Cloeotis, Coelops e Anthops.
Cloeops é o grupo irmão de Triaenops-Rhinolophus-Rhinonicteris.
Coelops e Anthops provavelmente estão relacionados filogeneticamente a Aselliscus.
Os Rhinolophoidea tiveram sua evolução praticamente restrita às regiões quentes do velho mundo (com pequena penetração na zona temperada). São divididos em 3 subgrupos (famílias): Megadermatidae, Nycteridae e Rhinolophidae. A família Megadermatidae é a mais conservadora com relação ao tamanho, características gerais e comportamento. As outras 2 famílias apresentam modificações significativas como redução do tamanho e outras características pedomórficas, como a redução do olho e modificações na folha nasal. A família Nycteridae conservou as grandes orelhas dos Megadermatidae, mas sofreu uma drástica redução no tamanho do olho e grande modificação na folha nasal e narinas, uma pedomorfose comparável a certas anomalias da nossa espécie como a mal formação conhecida como "lábio leporino" (associada com ausência de septo nasal e separação dos incisivos numa unidade central). A família Rhinolophidae sofreu redução no tamanho das orelhas, na folha nasal e olhos (mais evidente no gênero Rhinolophus). Essa grande diferenciação de Rhinolophus gerou a separação desse grupo numa família isolada (visão gradista que sempre separa tudo que é diferente e que parece não ter fim) criando consequentemente uma família parafilética, Hipposideridae.
Os Megadermatidae asiáticos são mais conservadores que os africanos. Uma indicação de centro de origem (ou centro de dispersão). A conquista de novos ambientes geralmente vem acompanhada de uma adaptação a esses ambientes.
O gênero Nycteris apresenta maior diversidade na África e Nycteris arge (espécie africana) é considerada basal na evolução desse gênero. Isso aponta a África como centro de origem de Nycteridae.
Rhinolophidae sendo o grupo mais derivado deve ter se originado (como já comentei) na região de quebra da distribuição das florestas tropicais. Rhinolophidae é também o grupo que invadiu a região temperada, portanto deve ter se originado entre o deserto do Saara e o Rajastão. Assim como os felinos e outros grupos de origem asiática, os Chiroptera devem ter se dispersado através das cadeias montanhosas, e sua diferenciação durante uma fase climática mais seca deve estar relacionada às cadeias montanhosas, inclusive á fase de adaptação à vida nas cavernas
The Rhinolophoidea had its evolution largely confined to warm regions of the Old World (with little penetration in the temperate zone). They are divided into 3 subgroups ( families ) : Megadermatidae , Nycteridae and Rhinolophidae.
The family Megadermatidae is the most conservative with respect to the size, general characteristics and behavior. The other 2 families show significant changes as a reduction in size and other paedomorphic characteristics such as eye reduction and changes in nasal leaf. The family Nycteridae retained big ears of the Megadermatidae, but suffered a drastic reduction in eye size and larger change in nasal leaf and nostrils, a paedomorphosis comparable to certain anomalies of our species, as the malformation known as "harelip" (associated with absence of nasal septum and separation of the incisors in a central unit). The Rhinolophidae family suffered reduction in the size of the ears, in nasal leaf and eyes (most evident in the genus Rhinolophus). This great difference in Rhinolophus generated the split of this group in a separate family (phenetic vision that always separates everything is different and seems to have no end) thus creating a paraphyletic family Hipposideridae.
EVOLUTION of the nasal leaf in RHINOLOPHIDAE

HOMOLOGIES in RHINOLOPHIDAE - a classical poster. This publication points to the future.
Pay attention on VIBRISSAE. It's very important to understand de homologies.
1 - Aselliscus presents a more conservative nasal leaf. The structure of the trident resembles the tip of the nasal leaf of Megaderma and nostrils are less exposed than in other genres. In my view it is possible to understand the process that led to the reduction of a type of nasal leaf such as in Megaderma, if we compare the figures of M. spasma of the plaque, with the schema that represents Aselliscus. There was an invagination in the central region of the nasal leaf, accompanied by a shortening.
figures of Paratriaenops, Triaenops, Rhinonicteris and Cloeotis from Hill, 1982.
figure of Hipposideros from Jenkins, P. D., and J. E. Hill. 1981.
figure of Asellia from DeBlase, A.F. 1980.
figure of Asellicus made by myself based on photos.
figure of Rhinolophus modified from Wu, Yi, Motokawa, M. and Harada, M. 2008.
References:
Benda, P. and Vallo, P.. 2009. Taxonomic revision of the genus Triaenops (Chiroptera: Hipposideridae) with description of a new species from southern Arabia and definitions of a new genus and tribe. Folia Zoologica 58(Monograph 1):1–45.
DeBlase, A.F. 1980. The bats of Iran: Systematics, distribution, ecology. Field Museum of Natural History (Chicago, Ill).
Jenkins, P. D., and J. E. Hill. 1981. The status of Hipposideros galeritus Cantor 1846 and Hipposideros cervinus (Gould 1854) (Chiroptera: Hipposideridae)., Zoology Series, 41(5): 279-294.
Hill, J. E. 1982. A review of the leaf-nosed bats Rhinonycteris, Cloeotis and Triaenops (Chiroptera: Hipposideridae). Bonner Zoologische Beiträge 33:165–186.
Wu, Yi, Motokawa, M. and Harada, M. 2008. A New Species of Horseshoe Bat of the Genus Rhinolophus from China (Chiroptera: Rhinolophidae). Zoological Science 25(4):438-443. 2008
1- Aselliscus apresenta uma folha nasal mais conservadora. A estrutura do tridente se assemelha à extremidade da folha nasal de Megaderma e as narinas estão menos expostas que nos demais gêneros. Ao meu ver é possível entender o processo que levou à redução de um tipo de folha nasal como o de Megaderma, se compararmos as figuras de M. spasma da prancha com o esquema que representa Aselliscus. Houve uma invaginação da região central da folha nasal, acompanhada de um encurtamento.
Rhinolophidae Evolution 2

Usando informação da literatura sobre a musculatura associada ao hióde nos Rhinolophidae, duas espécies de Nycteris (consideradas como basais) como grupo irmão imediato e Emballonuridae como um representante dos Vespertiliformes. Concluí que os Rhinolophidae apresentam pelo menos duas características sinapomórficas e Triaenops + Rhinolophus uma característica, nessa região do sistema muscular.
Quero deixar claro que esses caracteres não são simplesmente pequenos detalhes insignificantes. Mas representam transformações significativas na anatomia desses animais, pois alteram bastantes suas proporções corpóreas, certamente com implicações fisiológicas significativas. Provavelmente estão associados a emissão de sons, pois Hipposideros e Rhinolophus possuem traquéias extremamente robustas, com um diâmetro três vezes maior que no grupo-irmão. Triaenops apresenta uma traqueia mais conservadora, semelhante a Nycteris, mas ainda assim um pouco mais robusta. Portanto deve ter ocorrido desenvolvimentos independentes em Hipposideros e Rhinolophus.
Uma traqueia com maior diâmetro deveria emitir sons de baixa frequência, foi exatamente o que se detectou em Rhinolophus : 77- 81 kHz - (Schober e Grimmberger, 1997). A som é emitido a partir do nariz, e não pela boca (Nowak, 1994).
O desenvolvimento dessa musculatura associada a uma traqueia robusta é compreensível, pois é necessária mais energia para produzir tais sons. Esse emissor de som está associado a uma folha nasal direcionadora e a uma câmara nasal no crânio, aparentemente os morcegos menores possuem o crânio mais inflado no focinho.
Using literature data about the musculature associated with the hyoid in the Rhinolophidae, two species of Nycteris (considered as basal) as immediate sister group and Emballonuridae as a representative of Vespertiliformes.
I concluded that the Rhinolophidae have at least two synapomorphic features and Triaenops + Rhinolophus one feature in this region of the muscular system. I want to make clear that these characters are not just small insignificant details. But represent significant changes in the anatomy of these animals, because their body proportions change quite, certainly with significant physiological implications. They are probably associated with the emission of sounds, as Hipposideros and Rhinolophus have extremely robust tracheas with a diameter three times larger than the sister group. Triaenops presents a more conservative trachea, similar to Nycteris, but still somewhat more robust. Therefore must have occurred independent developments in Hipposideros and Rhinolophus.
A trachea with larger diameter should give low-frequency sounds, it was exactly what was detected in Rhinolophus: 77- 81 kHz - (Grimmberger and Schober, 1997). The sound is emitted from the nose, not the mouth (Nowak, 1994).
The development of these muscles associated with a robust trachea is understandable, since more energy is required to produce such sounds.
This sound transmitter is associated with a guiding nasal leaf and to a nasal chamber in the skull, apparently the smallest bats have the most inflated skull on the muzzle.
References:
Griffiths, T. A. 1994. Phylogenetic Systematics of Slit-faced bats (Chiroptera, Nycteridae). based on hyoid and other morphology. Americam Museum Novitates, 3090, 1-17.
Griffiths, Thomas Alan.; Koopman, Karl F.; Starrett, Andrew, 1930. The systematic relationship of Emballonura nigrescens to other species of Emballonura and to Coleura (Chiroptera, Emballonuridae). American Museum Novitates ; no. 2996.
Nowak, R. 1994. Walker's Bats of the World. Baltimore, Maryland: The Johns Hopkins University Press.
Smith, Andrea L.1990. Systematic Relationships of Rhinolophid Bats: Based on Hyoid Morphology. Illinois Wesleyan University.
Schober, W., E. Grimmberger. 1997. The Bats of Europe and North America. New Jersey: TFH Publications, Inc.
Hipposideros diadema and H. cineraceus - Mandible Comparison

Comparação das mandíbulas de Hipposideros diadema e Hipposideros cineraceus.
A polarização dos caracteres deve obedecer a ordem de tamanho.
Quanto menor for uma espécie mais derivada. Devemos lembrar que redução de tamanho nesse caso, e na maioria dos gêneros de morcegos, obedece as regras da pedomorfose. Veja aqui os ângulos e proporções da mandíbula.
A comparação das mandíbulas e a diferença de tamanho existente entre essas espécies, implicando em diferentes ecologias, já é suficiente para se determinar que Hipposideros diadema é sem dúvida uma espécie conservadora e H. cineraceus derivada e afastada do padrão ancestral em vários aspectos. Outra evidência forte e correlacionada é o arco zigomático. Uma mandíbula robusta, com processos mais robustos, costuma estar associada a um arco zigomático compatível e a um crânio com cristas temporais, sagital e ocipitais mais desenvolvidas, como neste caso. Mesmo assim à medida que vamos comparando todas as espécies, mais caracteres deverão corroborar ou afastar essa hipótese.
Comparison of the mandibles in Hipposideros diadema and Hipposideros cineraceus.
The polarization of the characters must obey the order of size.
The smaller is a species most derived. We must remember that size reduction in this case, and in most genera of bats, obeys the rules of paedomorphosis. See the angles and proportions of the mandible.
The comparison of the mandibles and the difference in size between these species, resulting in different ecologies, is enough to determine that Hipposideros diadema is undoubtedly a conservative species and H. cineraceus derived and away from the ancestral pattern in several respects.
The zygomatic arch is another strong and correlated evidence. A robust mandible, presenting more robust processes, is usually be associated with a compatible zygomatic arch and a skull with temporal, occipital and sagittal crests more developed, as in this case. Anyway as we get compared all species, more characters should corroborate or dismiss this hypothesis.
-These photos of the Hipposideros cineraceus skulls was published in:
A Taxonomic Review of Hipposideros halophyllus, with Additional Information on H. ater and H. cineraceus (Chiroptera: Hipposideridae) from Thailand and Myanmar
Bounsavane Douangboubpha, Sara Bumrungsri, Pipat Soisook, Susan W. Murray, Sébastien J. Puechmaille, Chutamas Satasook, Si Si Hla Bu, David L. Harrison, Paul J. J. Bates.
Journal: Acta Chiropterologica - ACTA CHIROPT , vol. 12, no. 1, pp. 29-50, 2010
-Mandible photo of Hipposideros diadema from : Animal Diversity Web - University of Michigan.
Hill's Revision of Hipposideros

My view is based on my recent publications on FLICKR, that is my official vehicle of communication with the "BAT COMMUNITY" !
Beginning with the study of nose-leaf and also by studying the skull, having the MANDIBLE as a focus in search for understanding the evolution of the genre, I started this new endeavor. As I have mentioned numerous times ....
The mandible is directly related with the zygomatic arch, the crests of the skull and obviously with the premaxilla and maxilla.
In summary The MANDIBLE reflects the ecology, physiology, behavior ..... and this necessarily relates to the environment and consequently the BIOGEOGRAPHY.
Without a reconstruction of the ancestor is difficult to polarize the internal characters. For this we have to keep in mind the size gradient and variation of characters within the genre. The syntheses of Tate and Hill give us this initial overview to begin the revision.
The study of external groups and their degree of relationship is essential in the polarization of characters. So I began the posts from an overview of all Rhinolophidae.
But the genus Hipposideros is the most important in understanding the Rhinolophidae, being basal, being poorly defined and cover a large number of species.
Começando pelo estudo das folhas nasais e também pelo estudo do crânio, tendo a mandíbula como foco em busca da compreensão da evolução do gênero, iniciei esse novo empreendimento. Como já citei inúmeras vezes....
A mandíbula se relaciona diretamente ao arco zigomático, às cristas da caixa craniana e obviamente às pré-maxilas e maxilares.
Em síntese A MANDÍBULA reflete a ecologia, a fisiologia, o comportamento.....e isso se relaciona necessariamente ao ambiente e consequentemente à BIOGEOGRAFIA.
Sem uma reconstrução do ancestral fica difícil polarizar os caracteres internos. Para isso temos que ter em mente o gradiente de tamanho e a variação dos caracteres dentro do gênero. As sínteses de Tate e Hill nos fornecem esse panorama inicial para começar o trabalho de revisão.
O estudo dos grupos externos e o seu grau de relacionamento é indispensável na polarização dos caracteres. Por isso iniciei as postagens a partir de uma visão geral de todos os Rhinolophidae.
Mas o gênero Hipposideros é o mais importante na compreensão dos Rhinolophidae, por ser basal, por ser mal definido e abranger um grande número de espécies.
References:
Tate, G. H. H. (1941). A review of the genus Hipposideros with special reference to Indo-Australian species. New York: American Museum of Natural History.
Hill, J.E. 1963. A revision of the genus Hipposideros. Bulletin of the British Museum (Natural History), Zoology Series, 11:1-129.

Eu construí esse cladograma parcial, justamente para comparar com a filogenia proposta por Hill, 1963.
Nós podemos identificar perfeitamente o ramo basal colocado à direita, na filogenia proposta por ele. Nesse ramo ele mostra duas dicotomias: 1- armiger-pratti e 2- diadema-speoris.
Escolhi as espécies que dão seus nomes aos grupos para facilitar a compreensão.
A dicotomia armiger-pratti se justifica, pois esses grupos (aqui representados por duas espécies) compartilham um conjunto de caracteres associados a uma folha nasal bastante modificada e complexa.
Hipposideros pratti apresenta uma folha nasal como em nenhum outro grupo de Hipposideros. Nessa espécie existe uma estrutura adicional extravagante, que se desenvolveu por trás da folha nasal básica, exatamente atrás do elemento chamado de folha nasal posterior. Observe que essa estrutura quando vista de frente oculta completamente a visão dos olhos.
Em H.armiger podemos identificar com facilidade, um desenvolvimento especial por trás da folha nasal posterior, que representa um estágio anterior à formação da estrutura extravagante da folha nasal de H. pratti. Portanto dois estados de uma sinapomorfia bastante significativa.
Na figura acima à esquerda, temos a representação da segunda dicotomia proposta por Hill (1963), "diadema-speoris".
Aparentemente essa dicotomia não se justifica (pelo menos neste estágio da análise), ou seja, não consigo identificar nenhuma mudança significativa compartilhada por esse conjunto, além das simplesiomorfias.
O posicionamento de Hipposideros bicolor (representando seu grupo) logo acima de H. speoris está realçando a principal diferença existente entre a filogenia proposta por Hill e a minha visão da evolução desse gênero. H. speoris possui um conjunto de caracteres associados à diminuição de tamanho (por pedomorfose) que culminam no grupo H. bicolor, que na visão de Hill seria basal e conservador.
Na extremidade direita da figura, temos uma comparação das formas das mandíbulas de 3 grupos: diadema, armiger e bicolor. Observa-se claramente uma diminuição pedomórfica do tamanho, com redução dos processos coronoíde e angular, já ressaltada numa figura anterior desse álbum, mas colocada aqui para permitir uma integração das ideias.
Essa integração da análise do crânio, comparada à análise dos caracteres externos da cabeça, corrobora as relações por mim apresentadas para esses grupos de espécies, previamente propostos por Tate (1941) e Hill (1963).
References:
Tate, G. H. H. (1941). A review of the genus Hipposideros with special reference to Indo-Australian species. New York: American Museum of Natural History.
Hill, J.E. 1963. A revision of the genus Hipposideros. Bulletin of the British Museum (Natural History), Zoology Series, 11:1-129.
Rhinolophidae Skull Comparation - ventral view.

Uma visão atualizada da evolução dos RHINOLOPHIDAE, com um rearranjo das subfamílias Hipposiderinae e Rhinolophinae.
Pela primeira vez uma definição precisa e evolutiva dos Hipposiderinae e dos Rhinolophinae.
Os HIPPOSIDERINAE com apenas 2 gêneros; Hipposideros e Asellia, os demais gêneros estão em RHINOLOPHINAE. Apresento também as relações de alguns subgrupos de Hipposideros.
With regard to the skull....apparently Aselliscus is a Hipposideros miniature..... that appeared in the early diversification of Rhinolophidae.
Just compare Aselliscus with Hipposideros pygmaeus, this species is like a revival of Aselliscus within the evolution of Hipposideros.
Still with regard to the skull......while in the evolution of Hipposideros happened a decrease in size, in the Rhinolophinae the opposite occurred.............. increasing size ( starting from a paedomorphic form but with increase in size, preserving the proportions).
Rhinolophidae Synapomorphies


1 - Uma importante sinapomorfia dos Rhinolophidae é a redução das bacias hipoconais dos molares superiores.
A Infraordem dos Noctiliformes se caracteriza por apresentar amplas bacias hipoconais nos molares superiores, mas estão reduzidas nos Rhinolophidae.
2 - As pré-maxilas dos Rhinolophidae são peculiares, estão restritas à região palatal e cada uma delas possui apenas um incisivo. Essa é também uma característica distintiva desse grupo de RHINOLOPHOIDEA.
Já vimos anteriormente mais duas sinapomorfias relacionadas a caracteres externos:
3 - Redução das orelhas.
4 - Perda do TRAGUS.
5 - Musculatura ver acima: Rhinolophidae Evolution 2
6 - Folha nasal ver acima: Evolution of the nasal leaf in Rhinolophidae
Podemos também observar nessa figura, as diferenças existentes entre as regiões posteriores dos palatos dos Rhinolophinae e dos Hipposiderinae. Os Hipposiderinae (Asellia e Hipposideros) possuem uma redução no tamanho dos terceiros molares superiores, acompanhada por uma reentrância palatal, bastante evidente no gênero Asellia (acompanhada por uma redução proporcional do molar). Essas 2 características são também sinapomorfias dos Hipposiderinae, são exclusivas desse grupo. Veja a comparação dos molares numerados.
-Infraordem : Vespertiliformes
A Infraordem: Vespertiliformes é formada por 2 superfamílias :
1. Vespertilionoidea [ Myotidae, Plecotidae, Otonycteridae, Histiotidae, Laephotidae, Antrozoidae, Nyctophilidae, Vespertilionidae, Emballonuridae, Rhinopomatidae, Craseonycteridae e Molossidae].
2. Nataloidea [ Myzopodidae, Thyropteridae, Natalidae e Furipteridae ].
Esses morcegos derivaram a partir de um ancestral comum, que sofreu redução no tamanho do corpo através de pedomorfose. Isso implicou em redução extrema das pré-maxilas resultando no aparecimento de um diástema entre os incisivos. Esse diástema é progressivo e tem sua expressão máxima nos Vespertilionidae. Dentre os Nataloidea só mesmo os Thyropteridae apresentam um diástema comparável aos Vespertilionidae mais basais, nos demais a separação dos incisivos pode ser mais sutíl.
Esses morcegos se destacaram bem na base da evolução dos Microchiroptera, pois tanto na base dos Vespertilionoidea como nos Nataloidea a dentição é a mais completa dentre os Chiroptera, semelhante aquela de Onychonycteris [ morcego fóssil do início do Eoceno ].
A redução do tamanho do corpo implicou também numa nítida redução do encéfalo, particularmente do telencéfalo, deixando o cerebelo proporcionalmente maior que na outra Infraordem dos Microchiroptera, os Noctiliformes.
Essa Infraordem preservou não apenas o número de dentes [38 no total], mas também a forma dos molares que na outra Infraordem sofreu modificação. No entanto os incisivos se mostram mais reduzidos em função da transformação da região anterior do focinho e aparecimento do diástema.
O rinário, que na outra Infraordem pode apresentar folha nasal ou narinas mais ou menos tubulares, nesses apresenta-se mais conservador, com pouca transformação além de uma pequena redução em virtude da diminuição do tamanho do corpo.
Os Vespertilionoidea caracterizam-se por apresentar um diástema mais pronunciado comparativamente, e essa é a marca mais evidente nos crânios.
Os Nataloidea caracterizam-se por uma evolução que partiu de uma adaptação muito especial que aparece em duas famílias e que depois transformou-se mais ainda, nas outras duas famílias restante. Eles adquiriram ventosas nos polegares e nos calcanhares, pelas quais se fixam a superfícies lisas geralmente folhas. As outras 2 famílias perderam essas ventosas durante o processo evolutivo do grupo, mas em função dessa perda o polegar que havia se transformado em ventosa acabou se reduzindo.
Dentre os Nataloidea, os Myzopodidae são os mais conservadores, apresentam além das ventosas, rinários mais conservadores (ainda que modificados, com narinas arredondadas) e orelhas maiores que lembram as orelhas do grupo irmão os Vespertilionoidea.
As outras 3 famílias possuem orelhas menores e rinários mais modificados.
Os crânios apresentam a região inter-orbital mais dobrada, provavelmente devido a redução do tamanho dos olhos. Esse dobramento atinge um grau extremo em Furipteridae, formando um ângulo de quase 90°. Todas essas características podem se vistas na figura acima.
NATALOIDEA
VESPERTILIONOIDEA
_________________________________________________________________________________________
BLUMENBACH, Johann Friedrich: Handbuch der Naturgeschichte. Bd. 1. Göttingen, 1779. LINK
ILLIGER, J. K. W. 1811. Prodromus systematis mammalium et avium additis terminis zoographicis utriusque classis, eorumque versione germanica. Sumptibus C. Salfeld, Berolini [Berlin]: [I]-XVIII, [1]-301, Errata et Omissa. LINK
________________________________________________________________________________________
 |
| Microcebus nos ajuda a pensar sobre como seria o ancestral dos primatas atuais. Porém a cauda teria escamas, como Tarsius. |
ARBOREA (Gomes, 2015) : Superorder ARCHONTA (Gregory, 1910), Primates (Linnaeus, 1758)
Em 1758, Carolus Linnaeus criou no seu SYSTEMA NATURAE, a ORDINES PRIMATES numerada como I, sendo o gênero HOMO também o primeiro da lista, isso é muito significativo. A sua diagnose da Ordem como todas daquela época era muito simples e eu a apresentarei aqui, a seguir, apenas como um registro histórico:

Dentes primores superiores IV. paralleli. Laniarii solitarii. Mammae pectorales, binae.
Palmae Manus sunt.
Brachia diducta claviculis, incessu tetrapodo valgo.
Scandunt arbores earumque gazas legunt.
 |
| Propositalmente temos um destaque em Hominidae (Superfamily Hominoidea), para evidenciar a parafilia, da artificial "Superfamily Cercopithecoidea". Vejam por exemplo, que Papionidae, Macacidae e Hominidae, possuem focinhos proeminentes, o que significa uma série de modificações a partir da forma ancestral. Isso permite mais espaço para os dentes, que são maiores, significa mudança na relação de forças que atuam no pescoço, com consequentes mudanças na musculatura. Também a olfação e a postura mudam. Essas famílias apresentam a região inter-orbital mais achatada e o espaço entre os olhos é mais curto, deixando os olhos mais juntos, numa cara alongada. |
Os gêneros da famila Hominidae, tanto os Hylobatinae como os Homininae, apresentam encéfalos muito derivados. Hylobates pode ser considerado como o mais simples se comparado aos demais, mas mesmo assim já representa um salto na evolução desse órgão, comparado aos grupos irmãos. Em complexidade poderia ser comparado aos babuínos, mas estes evoluíram numa outra linha, a partir de formas semelhantes a Cercocebus ou Lophocebus ! Existem boas evidências de que Papio é mais relacionado a Lophocebus, enquanto Mandrillus está mais relacionado a Cercocebus.
Gilbert, Christopher C. 2007. Craniomandibular morphology supporting the diphyletic origin of mangabeys and a new genus of the Cercocebus/Mandrillus clade, Procercocebus.

Tarsius é um Haplorhini, não possui um rinário como os primatas mais antigos nem tapetum lucidum e perdeu a função da enzima terminal que fabrica vitamina C. Portanto deveria estar junto com os Platyrrhini. Coerentemente Aotus não possui tapetum lucidum como Tarsius, pois essa característica foi perdida no ancestral de ambos. O argumento em favor da origem diurna de Aotus fica comprometido, pois Tarsius é noturno e não tem "tapetum lucidum". Podemos dizer com base em Tarsius e no seu grau de conservadorismo, que provavelmente o ancestral dos Platyrrhini era noturno. "Among primates a tapetum lucidum cellulosum is found in several prosimians, some of them nocturnally active, but also, for example, in the diurnal Lemur catta as well as in those diurnally active Indridae whose retinas have been studied. A tapetum is presumed to be missing in the unique nocturnal prosimian Tarsius and also in several diurnal or crepuscular Eulemur species (e.g., Eulemur macaco). It appears that there is no obvious correlation between the diarhythm of an animal species and its possession or lack of a tapetum. The tapetum is missing in the tree shrew genus Tupaia, but is present in all the Lorisidae and Galagidae... "Eyes and Eyesight." Primate Anatomy - Friderun Ankel-Simons, pg 375.
Em função desse novo arranjo das relações filogenéticas, a classificação dos Platyrrhini ficaria diferente:
FAMILY: Aotidae
FAMILY: Callitrichidae
FAMILY: Cebidae
subfamily: Callicebinae
subfamily: Pithecinae
subfamily: Alouattinae
subfamily: Cebinae
( Ateles, Brachyteles, Lagothrix, Oreonax, Cebus, Sapajus and Saimiri ).
A evolução dos Platyrrhini mostra que a redução do tamanho do corpo nos Callithrichidae e em Saimiri, levou a uma redução do número de circunvoluções tornando o encéfalo mais simplificado. Um processo evolutivo conhecido como pedomorfose deve ter agido nesse sentido. Para podermos entender esse processo temos que imaginar um organismo em desenvolvimento. Durante o desenvolvimento o corpo cresce como um fractal. Existem muitas sequências que trabalham simultaneamente e essas sequências se iniciam por "indução embrionária". Todo o desenvolvimento cessa em um determinado estágio embrionário que se assemelha a um estágio evolutivo ancestral. O aumento do tamanho do corpo está obviamente relacionado com a complexidade das circunvoluções. O número de circunvoluções não está exclusivamente ligado à inteligencia do animal, esse número está associado ao tamanho do corpo e ao número de integrações entre as diferentes partes do sistema nervoso: movimentos, sensibilidade e principalmente com a integração das diferentes áreas de ação neuronal. Elefantes, rinocerontes e baleias possuem um corpo grande e um grande número de circunvoluções no encéfalo. Se existem muitas áreas sensíveis devem existir muitas áreas de recepção e integração com o sistema, esse raciocínio é lógico.
 |
| Dois ramos de Platyrrhini sofreram pedomorfose: os Callitrichidae e os Cebinae [Cebus e Saimiri]. Na análise que venho fazendo desses primatas, tenho concluído que um ramo dos Cebinae: Cebus-Saimiri , sofreu um acúmulo de 2 processos seguidos de pedomorfose. Na figura acima vemos perfeitamente que existe uma nítida gradação quanto ao número de circunvoluções do encéfalo de Aotus até Ateles. Particularmente Saimiri, que pode ser visto na figura precedente, sofreu uma simplificação. Cebus no entanto parece perfeitamente encaixado na série de transformações, e mesmo que tenha sofrido simplificação, não foi o suficiente para mascarar o processo. Em Cebus e Saimiri temos uma perda gradual da condição "cauda preênsil": semi-preênsil em Cebus e totalmente ausente em Saimiri. Considero a condição não preênsil da cauda de Saimiri como uma evidência de perda gradual em Cebinae. |

LITERATURA
Gilbert, Christopher C. 2007. Craniomandibular morphology supporting the diphyletic origin of mangabeys and a new genus of the Cercocebus/Mandrillus clade, Procercocebus.
__________________________________________________________________________________________

ARBOREA (Gomes, 2015) : Superorder ARCHONTA (Gregory, 1910) Scandentia (Wagner, 1855)
Os SCANDENTIA foram assim batizados por Wagner, em 1855, como uma das 5 Famílias dos INSECTIVORA. Como sempre acontecia com as descrições feitas pelos europeus, eles foram comparados aos esquilos, ou seja, uma referência a um animal europeu. O nome SCANDENTIA vem do latim scandens, que significa "aquele que escala" (significando portanto: Os Insetívoros Escaladores). Esses animais são internacionalmente conhecidos como Tupaias, que vem da palavra malaia tupai, usada regionalmente para se referir a um esquilo ou a um musaranho. Wagner (1855: 524) os descreveu em latim e alemão (sua língua-mãe) da seguinte forma:
II. Familie.
Scandentia. Spitzhörnchen
Habitus sciureus at rostro valde elongato attenuato, dentes canini sejuncti, annulus orbitalis osseus clausus.
Leben gleich Eichhörnchen auf Bäumen, sind auf Südasten beschränkt, und bei ihnen kann man beireits ächte Eckzähne unterscheiden, die, wenn auch nicht ihrer Fucktion nach, doch wegen ihrer Form, aberünckten Stellung einwurtzeligen beschaffenheit als solch erklärt werden können. Sie nähren sich von Insekten, deren Larven und Eiern, so wie auch von saftigen Früchten.
Scandentia. Esquilos (de focinho) pontudo
O aspecto lembra o de um esquilo, com focinho bastante alongado e estreito, dentes caninos separados e anel orbital fechado por osso.
Vivem nas árvores como os esquilos, estão restritos ao Sudeste (da Ásia), e neles podemos distinguir oito caninos, que, se não na sua função, podem ser explicados como tal pela sua forma, posição curvada e natureza unirradicular. Alimentam-se de insetos, suas larvas e ovos, além de frutas suculentas.
_____________________________________________________________________________
Tupaia glis foi descrita em 1820 por Diard & Duvaucel como Sorex Glis. O animal foi num primeiro momento, confundido com um esquilo. Porém logo notaram que suas características eram de um musaranho, mais exatamente, de um Sorex. Entretanto, baseados em suas características anatômicas, reconheceram que o animal poderia ser considerado como pertencente a uma nova subdivisão.
página 472 On the Sorex Glis
Notificação, - Sobre uma nova espécie de Sorex. - Sorex Glis (D.D.) (Diard & Duvaucel)
Quando a investigação em história natural não tinha, por assim dizer, outro objectivo senão a acumulação de espécies e a distinção de formas externas; a descoberta de um pequeno animal que não teria sido notável, nem
473
pela sua forma, nem pela sua cor, nem pelos seus hábitos, não teria interessado muito aos naturalistas: mas hoje que a ciência quer sobretudo alargar o seu domínio, das observações anatômicas, e assim assegurar a sua marcha sobre caracteres invariáveis, a coisa que lhe parece mais preciosa já não é aquela que mais se distingue dos outros, pela riqueza do seu adorno, ou pela singularidade das suas proporções, mas sim aquela que pode fornecer o maior número de fatos para a confirmação ou a modificação das leis orgânicas que foram reconhecidas.
Hoje, finalmente, que o bisturi examinador provou que a natureza enterrou muitas vezes os seus mistérios mais admiráveis, nas formas mais vis e mais comuns, temos o direito de esperar que os naturalistas vejam com alegria o seu catálogo crescente na história de um nova espécie, que não só não tem nada de desagradável ou repugnante, mas que pelo contrário nos dá pela primeira vez o exemplo de um animal pequeno, dos mais graciosos, possuidor de todas as características genéricas, que pareciam estar reservadas exclusivamente a um poucos seres deformados ou revoltantes.
DURANTE nossas estadas em Pulo Penang e Singapura, matamos várias vezes na floresta um pequeno quadrúpede, que primeiro confundimos com um esquilo; mas que logo reconhecemos, ao examiná-lo, pertencer à família dos Insetívoros: só a forma alongada do seu focinho poderia ter-nos feito suspeitar que não se tratava de um roedor: porque, como acabamos de dizer, por todas as outras proporções do seu corpo, pelo tamanho, pelas orelhas raspadas, cobertas por pelos muito curtos, inteiramente moldadas como as de um humano, e principalmente pela disposição emplumada dos pelos da cauda, se assemelhava perfeitamente a uma pequena
Vol. XIV. 6E
474
espécie de esquilo, que encontramos a cada passo nas matas de Singapura: além disso, a sua cor não é notável; é marrom avermelhado na parte superior, misturado com fulvo e preto, e abaixo é cinza esbranquiçado uniforme; mas o que nos parece digno de nota é o tom rosado da pele deste lindo animal, que aparece principalmente ao redor dos olhos e lábios.
Se o focinho alongado e as patas pentadáctilas deste animal tornavam fácil reconhecer que pertencia à família dos Sorex, a singularidade da sua forma também poderia naturalmente levar à suposição de que não pertencia a nenhum dos gêneros que contém; e que é de fato, o que foi confirmado pelo número e disposição de seus dentes.
A mandíbula superior é armada com 4 incisivos, aproximadamente cilíndricos, não longos, ligeiramente desgastados em bisel e espaçados: dentes e molares em número de 5 e eriçados com pontas cônicas, é uma fileira isolada, aproximadamente do mesmo comprimento.
No maxilar inferior, ao contrário, existem 6 incisivos estreitos, voltados para a frente, dos quais os quatro intermediários são muito longos. O canino também é mais alongado que o anterior, tem atrás dele um pequeno molar falso, depois uma fileira de 4 molares tricúspides.
A estas particularidades na forma e nos dentes do nosso animal, se somarmos a presença de um pequeno ceco na origem dos Intestinos, um ceco que nenhum dos Sorex ainda apresentou, teremos certamente todo o direito possível de tomá-lo como o tipo de uma nova subdivisão: nós
475
vamos atribuir-lhe o nome (Sorex Glis) que dá tanto a ideia da sua forma externa como da sua verdadeira natureza.
FINALMENTE para finalizar a história desse verdadeiro Sorex, disfarçado com aspecto de esquilo, ele tem olhos grandes, 4 tetas ventrais, língua comprida, estômago simples e um tubo intestinal dobrado 7 vezes sobre si mesmo, e seguido como já dissemos, do pequeno ceco.
Este pequeno animal alimenta-se de insectos e principalmente de larvas que procura nos troncos das árvores velhas, e até pousa sob restos de folhas: raramente o encontramos, e sempre em locais isolados; Porém, parece que não é de natureza muito selvagem, porque quando estávamos em Penang, uma pessoa daquele lugar tinha um muito manso, que comia numa gaiola como um esquilo.
Diard & Duvaucel
 |
| Tupaia glis foi descrita em 1820 por Diard & Duvaucel como Sorex Glis. |
_____________________________________________________________________________
O gênero Tupaia, só foi proposto em 1821 por Thomas Stamford Raffles (um ano depois de sua descoberta por Diard & Duvaucel). Ele propôs esse gênero quando descreveu duas novas espécies de tupaias: T. ferruginea e T. tana, porém não fez nenhuma referência a Sorex glis de Diard & Duvaucel. Raffles criou o gênero porque coletou e identificou dois animais bem diferentes entre si e também por não se encaixarem em nenhum outro grupo já existente. Abaixo forneço a sua definição do gênero e das duas espécies de tupaias:
"TUPAIA. Caracteres genéricos. Focinho alongado. Dentes, Incisivos acima de 4 afastados. abaixo de 6 (ou 4?) inclinados para frente, os do meio aproximam-se. Caninos dois de cada lado, acima e abaixo, afastados. Molares, três de cada lado, acima e abaixo. Pés com cinco dedos. Aparência e cauda de um esquilo".
TUPAI Malaio توڤاي
Este pequeno animal singular foi observado pela primeira vez domesticado na casa de um cavalheiro em Penang, e depois encontrado selvagem em Cingapura e nas florestas perto de Bencoolen, onde vive das frutas; dos Kayo Gadis, etc. Possui cauda e aparência geral de esquilo, unida à cabeça alongada e dentição de Sorex. Difere ainda deste último gênero por ser um animal animado e brincalhão, com olhos grandes adequados à luz do dia e por não ser noturno ou subterrâneo em seus hábitos.
A cabeça prolonga-se num focinho cônico; a mandíbula superior e o lábio não se projetam muito além dos inferiores. Incisivos quatro acima, bastante remotos e pontiagudos, seis abaixo, os médios menores, os laterais inclinados para frente. Caninos quatro em cada mandíbula, distantes dos incisivos superiores. Seis molares cônicos em cada mandíbula. Olhos grandes e brilhantes. Orelhas arredondadas. Cor marrom ferrugem no dorso e nas laterais, esbranquiçada no ventre; cauda marrom-acinzentada, sendo os pelos uma mistura de preto e branco. Comprimento do corpo cerca de seis ou oito de uma Coleção Zoológica feita em Sumatra. 257 oito polegadas. Cauda não tão longa, peluda como a de um esquilo, mas mais redonda. Cinco dedos em todos os pés; os três do meio mais longos; todos equipados com garras curvas.
Esses animais são tão mansos e alegres quanto os esquilos. O domesticado acima mencionado andava em perfeita liberdade, percorria livremente toda a casa, e nunca deixava de se apresentar à mesa do café da manhã e do jantar, onde comia frutas e leite.
Esta espécie se assemelha muito à anterior, mas é maior, o comprimento do corpo é de 23 centímetros e o da cauda, de cerca de sete. A cor acima é quase a mesma, mas as partes inferiores são de um vermelho ferruginoso. A cauda desta espécie é plana como a de um esquilo e é de cor avermelhada.
No único exemplar já obtido deste último, existem apenas quatro incisivos na mandíbula inferior, mas em todos os outros aspectos os dentes concordam com a espécie anterior. Como o espécime foi preparado por um nativo antes de chegar às minhas mãos, será necessário um exame mais aprofundado para verificar se essa diferença não pode ter sido o efeito de um acidente na preparação.
Esses animais são geralmente conhecidos pelos nativos pelo nome de Tupai tana; e diz-se que são sempre encontrados no solo ou perto dele. Não é impossível que consigamos encontrar outras variedades; de qualquer forma, os dois observados são suficientemente distintos para formar espécies separadas e, presumo, para justificar a adoção de um novo gênero: certamente não pertencerão a nenhum dos gêneros já observados.
 |
| Tupaia tana variety chrysura. Proceedings of the Zoological Society of London (1876) by John Gerrard Keulemans. |
_____________________________________________________________________________
 |
| Thomas Henry Huxley |
382 A ANATOMIA DOS ANIMAIS VERTEBRADOS
Assim como os Rodentia, os Insectivora possuem grande diversidade de hábitos; alguns Galeopitheci planam pelo ar à maneira dos esquilos voadores; alguns arbóreos, como os Tupayae; alguns terrestres e cursoriais, como a maioria da ordem. Alguns são nadadores; e alguns, como as toupeiras, são os mamíferos mais completamente fossoriais. Vejam a ordem : planadores, arbóreos, terrestres cursoriais, nadadores e fossoriais.
A forma mais aberrante dos Insectivora é o gênero Galeopithecus, essencialmente um insetívoro de hábito arbóreo e frugívoro, com membros muito longos e delgados. Estes estão ligados entre si, com os lados do pescoço e do corpo, e com a cauda, por uma grande prega do tegumento, que se chama patágio; e, ao contrário da membrana da asas dos morcegos, é peluda em ambos os lados e se estende entre os dedos dos pés. Com a ajuda desta grande expansão semelhante a um pára-quedas, o Galeopithecus consegue dar saltos flutuantes, de árvore em árvore, através de grandes distâncias. Quando em repouso, os galeopitecos suspende-se pelas patas dianteiras e traseiras, com o corpo e a cabeça pendurados para baixo; uma posição que às vezes é assumida pelos saguis entre os primatas.
Os membros anteriores são ligeiramente maiores que os membros posteriores. Existem quatro tetas axilares. O macho tem pênis pendente e bolsas escrotais inguinais. O pólex e o hálux são curtos e capazes de movimentos consideráveis em adução e abdução, mas não são oponíveis; e suas garras são como as dos outros dedos.
O forame occipital está na face posterior do crânio. A órbita está quase, mas não totalmente, cercada por ossos. O forame lacrimal está na órbita. O teto ósseo do palato é largo e sua margem posterior é espessada. Existe um forte processo pós-glenoidal curvo do escamoso, que se une ao mastoide, abaixo do meato auditivo, e restringe o movimento da mandíbula ao plano vertical. Um corte longitudinal do crânio mostra uma grande câmara olfativa que se projeta além daquela dos lobos cerebrais, e duas cristas longitudinais, na face interna destes últimos, provam que esses lobos devem possuir sulcos correspondentes. O plano do tentório [membrana que separa o cerebelo dos lóbulos ocipitais do cérebro] é quase vertical e as fossas floculares [relativas ao cerebelo] são muito profundas.
A ulna é muito delgada inferiormente, onde se torna anquilosada à extremidade distal do rádio, que contém o carpo. Quando os ílios estão horizontais, os acetábulos olham um pouco para cima e para trás, bem como para fora. A fíbula é completa. Como nas Preguiças e na maioria dos Primatas, o navicular e
GALEOPITHECUS 383
o cuboide giram prontamente sobre o astrágalo e o calcâneo, de modo que o planta pedis é virado para dentro.
A fórmula dentária é i 2.2 / 3.3 , c 1.1/1.1 , pm + m 5.5/5.5 = 34
O incisivo externo, na mandíbula inferior, tem raízes, uma peculiaridade que não ocorre em outros lugares. Os caninos de ambas as mandíbulas também possuem duas raízes, como em alguns outros Insectivora. Os incisivos inferiores têm presas únicas; e suas coroas são largas, planas e divididas por numerosas fissuras longitudinais profundas, ou pectinadas. O comprimento de todo o canal alimentar, da boca ao ânus, não é superior a seis vezes o comprimento do corpo. O ceco saculado é tão longo quanto o estômago e sua capacidade deve ser maior que a deste último órgão.
Galeopithecus foi, uma vez, colocado entre os lêmures e, em outra, entre os morcegos. Mas as semelhanças com os primeiros são gerais e superficiais, e as diferenças na forma do cérebro, na dentição, na estrutura dos membros e do crânio, excluem-no da ordem dos Primatas.
Galeopithecus concorda com os morcegos na disposição da cauda e na existência de um patágio dotado de “músculos” especiais. Além disso, numa ligeira obliquidade do acetábulo, como se vê no seu desenvolvimento extremo nos morcegos; na condição imperfeita da ulna; e na posição peitoral das tetas e do pênis pendente. Estas 2 últimas, entretanto, deve ser lembrado, também são características primáticas [plesiomórficas]. Finalmente, os incisivos inferiores pectinados de forma semelhante são encontrados nos gêneros de Cheiroptera, Diphylla e Desmodus.
Mas Galeopithecus difere completamente dos morcegos pela estrutura dos membros anteriores; pela posição dos membros posteriores e pela ausência de calcar: por apresentarem duas presas nos incisivos externos e nos caninos; e pela presença de um ceco.
Por outro lado, as peculiaridades do crânio e do cérebro são principalmente insetívoras, assim como o canino de duas presas; e não vejo razão para discordar da opinião do Prof. Peters de que Galeopithecus não pertence nem aos primatas, nem aos quirópteros, mas que é um Insectivora aberrante [quando Thomas Huxley trata Galeopithecus como um Insectivora aberrante, está querendo dizer que é o mais derivado desse agrupamento, o mais afastado dos demais, e deu um tratamento todo especial a ele, pelo fato dele ser muito pouco conhecido na época, e como já citei acima, em 1869, Alfred Russel Wallace o tratou como um animal curioso, ou seja: "estranho para um europeu"].
Com relação aos demais Insectivora, vale ressaltar que Macroscelides apresenta o rádio e a ulna anquilosados. As tupaias possuem um grande ceco. Chrysochloris possui glândulas mamárias peitorais; Centetes e as toupeiras têm o pênis pendente.
384 A ANATOMIA DOS ANIMAIS VERTEBRADOS
Os Tupayae são animais de pelo macio, cauda longa e amantes das árvores, com órbitas ósseas completas e um grande ceco, e são os Insectivora que mais se aproximam dos lêmures.
Os musaranhos (Sorices) quase se parecem com roedores externamente, sendo muito parecidos com pequenos ratos. O zigoma é imperfeito, a tíbia e a fíbula estão anquilosadas e os ossos púbicos não se encontram na sínfise. Existem dezesseis a vinte dentes na mandíbula superior e doze na mandíbula. Os caninos estão ausentes e há seis incisivos acima e quatro abaixo. Os incisivos inferiores internos são muito alongados e inclinados, e alguns dos dentes não raramente ficam anquilosados com as mandíbulas. Não há ceco, e glândulas almiscaradas peculiares às vezes se desenvolvem nas laterais do corpo.
As toupeiras (Talpinae) não têm orelhas externas e os olhos são rudimentares. Os membros anteriores são muito maiores que os posteriores e estão incluídos no tegumento até o carpo. A superfície palmar da mão larga está voltada para fora e para trás. O manúbrio do esterno é muito largo e sua superfície ventral dá origem a uma crista mediana robusta. A escápula é tão longa quanto o úmero e o rádio juntos. Ela é triangular e possui um processo acromial, mas nenhum coracoide distinto [Vejam que ele termina as comparações com os mamíferos mais completamente fossoriais: as toupeiras, os quais ele tratou, igualmente, como aberrantes (derivados). Porém num polo oposto a Galeopithecus].
-DERMOPTERA-SCANDENTIA-MACROSCELIDES-ECHINOSOREX-SORICIDAE-TALPIDAE-SOLENODON-
Para terminar, ressalto aqui o papel do texto de Huxley na nossa análise dos Scandentia: sua origem e história taxonômica. A Ordem INSECTIVORA sempre foi um "saco-de-gatos", ou seja, nela foram colocados aqueles mamíferos basais, que não cabiam nos grupos taxonômicos tratados como superiores (claramente definidos e relativamente homogêneos, ou com uma clara continuidade de caracteres derivados). Essa retrospectiva é muito importante, pois a Ordem Scandentia ainda poderá ser modificada no futuro, uma vez que os Tupaiidae e os Ptilocercidae não formam um grupo homogêneo, sendo talvez até parafiléticos.
LITERATURA CITADA
DIARD, P.M., DUVAUCEL, A. (1820) "Sur une nouvelle espèce de Sorex — Sorex Glis". Asiatick researches, or, Transactions of the society instituted in Bengal, for inquiring into the history and antiquities, the arts, sciences, and literature of Asia, Volume 14. Bengal Military Orphans Press, 1822

ARBOREA (Gomes, 2015) : Superorder FERAE (Linnaeus, 1766) Carnivora (Bowdich, 1821)


 |
| Comparison of Carnivoran and Creodont Carnassials |
________________________________________________________________________________________

ARBOREA (Gomes, 2015) : Superorder UNGULATA (Linnaeus, 1766) Perissodactyla (Owen, 1848)
________________________________________________________________________________________

ARBOREA (Gomes, 2015) : Superorder UNGULATA (Linnaeus, 1766) Artiodactyla (Owen, 1848)
________________________________________________________________________________________
________________________________________________________________________________________

TERRESTRIA (Gomes, 2015) : Superorder ZALAMBDODONTA (Gill, 1883)
Solenodontoidea (Gomes, 2015), Chrysochloroidea (Broom, 1915), Tenrecoidea (McDowell, 1958)
 |
| Necrolestes patagoniensis da Argentina é um fóssil conhecido da Patagônia, que certamente encontra entre os Chrysochloridae os seus mais próximos relativos. Sua distribuição geográfica pode ser explicada pela conexão que existiu durante o Cretáceo entre a Península Antártica e o sul da África (Cráton de Kaapvaal). Nesse caso os †Necrolestidae seria o grupo irmão imediato de Chrysochloridae, por compartilharem caracteres únicos, e devem ser considerados como parte dos ZALAMBDODONTA. |



- Salton, Justine A.; Szalay, Frederick S. "The Tarsal Complex of Afro-Malagasy Tenrecoidea: A Search for Phylogenetically Meaningful Characters" Journal of Mammalian Evolution (2004-06-01) 11: 73-104 , June 01, 2004.
- Justine A. Salton and Eric J. Sargis. "Evolutionary morphology of the Tenrecoidea (Mammalia) carpal complex"
Biological Journal of the Linnean Society, 2008, 93, 267–288. With 11 figures.
- Justine A. Salton and Eric J. Sargis. "Evolutionary Morphology of the Tenrecoidea (Mammalia) Forelimb Skeleton". "Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay", 51–71. Springer Science + Business Media B.V. 2008.
- Justine A. Salton and Eric J. Sargis. "Evolutionary Morphology of the Tenrecoidea (Mammalia) Hindlimb Skeleton". JOURNAL OF MORPHOLOGY 270:367–387 (2009).

TERRESTRIA (Gomes, 2015) : Superorder Hypoconodonta (Gomes, 2015) Erinaceoidea (Gill, 1872)
Erinaceidae (Bonaparte, 1838), Soricidae (Gray, 1821), Talpidae (Gray, 1825)

 TERRESTRIA (Gomes, 2015) : Superorder Hypoconodonta (Gomes, 2015) Caecoidea (Gomes, 2015)
TERRESTRIA (Gomes, 2015) : Superorder Hypoconodonta (Gomes, 2015) Caecoidea (Gomes, 2015) Tubulidentata (Huxley, 1872) Macroscelidea (Gill, 1872) Peanungulata (Simpson, 1945) Glires (Linnaeus, 1758)



%20and%20Agnolin%20and%20Chimento%20(2011).jpg)

.jpg)
%20FOOT.jpg)